
| 일 | 월 | 화 | 수 | 목 | 금 | 토 |
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | ||
| 6 | 7 | 8 | 9 | 10 | 11 | 12 |
| 13 | 14 | 15 | 16 | 17 | 18 | 19 |
| 20 | 21 | 22 | 23 | 24 | 25 | 26 |
| 27 | 28 | 29 | 30 |
- 시각처리
- criminal psychology
- 집단주의
- memory
- #정신분석
- 외측 슬상핵
- #산업및조직심리학
- collectivism
- Psychology
- 행동주의
- 빅토리아 시대
- 테스토스테론렉스
- ctm
- individualism
- persuation
- 연결주의
- #정신역동
- 강화
- sexdifference
- 심리학
- 도덕발달단계론
- 행동주의 치료
- behavior modification
- 따뜻함주의
- heartism
- 개인주의
- 켈트 매듭
- #크립키
- celtic knot
- 일주율
- Today
- Total
지식저장고
주의 연구 정리 본문
주의는 환경 속의 여러 자극중 하나의 자극에만 집중할수 있게 하는 인지적 처리기제를 말한다. 주의가 있기 때문에 우리는 수많은 환경 속의 잡음 속에서 우리가 원하는 정보만을 선택하여 들을 수 있다. 또한 주의를 활용한 연구는 시각과 지각, 다른 인지과학 이론의 발달을 이끌었으며, 현재도 많은 인지과학자들이 피험자들의 초점을 조절하여 인간 인지에 대한 탐구를 이어나가고 있다.
주의1
주의(attention)는 환경 속의 여러 자극 중 특정 자극들을 인지하고 반응하게 하는 심리적 기제를 말한다. 주의는 환경의 지각과 정보처리에 중요한 역할을 담당하는데, 환경에는 정말 수많은 자극이 있지만 인간은 그중에서 소수의 자극만 지각하는데, 어떤 자극이 지각될지를 주의가 결정한다. 연구에 따르면 주의란 주의가 기울여지는 자극 관련 뉴런에 weight를 부과해서 해당 뉴런의 반응을 증폭하는 것이며, 이는 독립적인 뇌의 패턴으로 관찰할 수 있다.2 또한 biased-competition model에 따르면 서로 경쟁하는 뉴런들 간에 특정 뉴런과 뉴런 활동을 두드러지게 하고 다른 뉴런은 억제하는게 주의이다.
세부적으로 주의는 4가지로 나눌 수 있는데, 그 내용은 아래와 같다. alertness/arousal(각성)은 가장 기초적인 형태의 주의로, 전반적인 뉴런들의 활동 수준을 말한다. 동물은 각성이 일정 수준 이상일때 외부를 지각하고 자극에 반응할 수 있으며, 때문에 잠을 잘 때나 피곤할때처럼 각성이 낮은 상태에서는 각성이 낮아진다. 코마(coma)는 각성이 부재하는 상황을 말하는데, 코마 상태에 있는 사람들은 외부의 어떠한 자극에도 반응하지 못한다. vigilance(sustained attention)는 각성을 일정 시간 이상 유지하는 능력으로, 특정한 대상을 지속적으로 의식하는 능력을 말한다. vigilance는 특정한 일을 계속 집중하면서 해야할 때 중요하다.
주의를 기울이는데 드는 인지적 자원인 주의력은 유한하기 때문에 사람들은 한번에 하나의 대상에만 주의를 기울일 수 있다. 이때 목표하는 대상에만 선택적으로 주의를 기울이는 기제를 선택적 주의(selective attention)라 한다. 인간이 환경을 지각하는 경우에는 선택적 주의를 기울이는 대상에 지속적으로 초점이 고정되기 때문에 아이트래커를 사용하여 선택적 주의가 기울여지는 대상을 알 수 있다.
선택적 주의는 그 근원에 따라 bottom-up과 top-down으로 나눌 수 있다.3 bottom-up attentional selection(stimulus-driven attention, 상향식 주의, 자극-주도적 주의)은 지각된 환경 자극이 너무 독특하거나 우리의 본성에 부합해서 우리의 주의를 끌어오는 경우이고, top-down attentional selection(goal-directed attention, 하향식 주의, 목적-지향적 주의)은 의도적으로 특정 대상에 선택적 주의를 기울이는 경우를 말한다.
하지만 초점을 기울이지 않은 경우에도 주의를 기울이는게 가능은 한데(covert shift of attention), 이는 무의식적이고 파편적으로 환경에 가해지는 divided attention이 있기 때문이다. divided attention은 선택적 주의가 기울여지지 않는 대상에도 주의를 기울이는 능력으로, 멀티태스킹은 divided attention과 관련되어 있다.
주의 연구에서 많이 사용되는 방법은 dichotic listening이다. dichotic listening을 사용하는 연구자들은 피험자에게 헤드폰을 씌우는데, 헤드폰은 양쪽에서 각기 다른 음이 들린다. 대개 한쪽이 주의가 가해진 자극이고 다른 한쪽은 주의가 가해지지 않는 자극으로, 연구자들은 피험자에게 특정 음이 들린 쪽의 소리에 집중하라고 지시해서 그쪽으로 선택적 주의가 향하도록 한다. dichotic listening 절차를 시행한 결과 피험자들은 무시된 쪽에서 나는 음성은 거의 알아듣지 못했으며, 다만 말하는 이의 성별과 목소리톤은 기억했다.
주의는 다른 인지기제와 달리 특화된 영역이 없으며, 어느 정보처리에서건 사용가능하다. 이런 특성으로 인해 주의는 지각과정에서 필수적인 역할을 담당한다. 수많은 환경 자극들 중에서 오직 주의의 대상이 된 자극만 지각될 수 있다. 물론 주의를 기울이지 않은 자극도 지각될 수 있기는 하다.4 하지만 그 양은 매우 제한되고, 다른 연구에서 재현되지 않았다.
지각에서 주의가 차지하는 역할은 보이지 않는 고릴라 현상으로 익히 알려져 있다. 보이지 않는 고릴라 현상은 한 실험에서 나왔는데5 이 실험에서 피험자들은 사람들이 농구를 하고 있는 영상을 보고 그들이 패스를 총 몇 번 하는지 세어달라고 지시받았다. 영상이 끝나자 피험자들은 꽤 정확하게 세었는데, 이때 연구자가 피험자에게 영상에서 중간에 지나간 고릴라를 봤냐고 질문했다. 실제로 영상 중간에 고릴라 옷을 입은 사람이 나와 중앙에서 가슴을 두드리고 지나갔지만, 피험자 대부분은 고릴라를 보지 못했다. 이는 피험자들이 패스 횟수에만 주의를 기울였기 때문에 고릴라가 주의에 포착되지 못해 지각되지 않았기 때문이다.
아이트래커를 사용한 연구에 따르면 인간이 환경을 지각할때 색, 방향(특히 수직/수평 방향), 대비 등 특정 요소가 선택적 주의를 유발하는 경향이 있다. 이는 선택적 주의가 상향식으로 유발될 수 있음을 보여준다. 한 실험에서는 화면에 오른쪽을 가리키는 화살표를 놓고 피험자에게 곧 나올 불빛을 보라고 지시했는데, 불빛이 오른쪽에 나온 경우 왼쪽에 나온 경우보다 RT가 빨랐다. 반면 사람들은 대상에 존재할거라고 예상하는 자극에 더 초점을 맞춘다. 부엌에서 식빵에 잼을 바르는 사람 그림을 보여주면 초점은 잼에서 버터 나이프, 빵, 다른 곳 순으로 움직인다. 이는 선택적 주의가 하향식으로 유발될 수도 있음을 보여준다. 선택적 주의는 다른 지각과정과 마찬가지로 상향식 처리와 하향식 처리를 같이 수행한다.
선술했듯이 인간의 주의력은 생각보다 작다. 보이지 않는 고릴라 현상이 발생하는 이유도 사람이 거기까지 주의를 기울일 만큼 주의력이 여유있지 않기 때문이다. 그래서 복잡한 자극이 빨리 지나가버리면 그 자극은 대개 주의에 포착되기 힘들다. 변화맹(change blindness)은 인간이 짧은 순간의 변화를 감지하지 못하는 현상으로, 사람은 오직 특정한(그리고 변화가 일어날) 지점에 주의를 기울인 경우에만 변화를 감지할 수 있었다. 주의력이 작기 때문에 사람들은 단서를 통해 주의를 기울여야 할 범위를 한정한다. 연구에 따르면 사람은 불빛이 자극이 나타날 사분면이 제시되면 주의를 미리 기울이지 않아도 불빛을 더 잘 지각했다.
최근까지 어떤 과제이건 사용되는 주의는 하나이며, 때문에 매우 한정적인 주의력을 잘 분배하여 사용해야 한다고 여겨졌다. 그러나 multiple-resource theory6에서는 주의력이 단일한 자원이 아니라 특정 프로세스에 사용되는 특정 형태의 주의력들로 구성되어 있으며, 그렇기 때문에 서로 다른 과제를 할 때는 주의력을 둘러싼 갈등이나 주의력 저하가 일어나지 않는다고 주장한다. 실제로 이 이론은 왜 2개의 시각 과제를 하는 것이 1개의 시각 과제와 1개의 청각 과제를 하는 것보다 어려운지 잘 설명한다.
biased competition model
biased-competition model7은 주의의 본질을 설명하는 이론이다. 이 이론에 따르면 본래 자극들은 서로를 억제하는데,8 주의는 그러한 억제를 제거해줌으로서 주의가 가해진 자극들이 모두 잘 표상되도록 하고9 주의가 가해지지 않은 자극들은 억제하면서 작동하는 것으로 보인다.10 이처럼 서로 경쟁적인 신경 활동들 중에서 특정 활동이 두드러지도록 만드는 편향이 주의라는게 biased competition model의 설명이다.

computational model은 이론을 더욱 발전시켜서 하향식 주의와 상향식 주의가 서로 경쟁한다고 제안한다. 이에 따르면 하향식 주의 시스템과 상향식 주의 시스템은 자신이 선호하는 자극이 가장 큰 주의를 끌도록 서로 경쟁하며, 여기서 이긴 쪽에 선택적 주의가 가해진다. 그래서 특정 자극이 활성화되는 정도는 그것의 salience나 하향식 주의가 가해지는 정도에 영향을 받는다.11 이 모델에서는 stroop task도 상향식 주의 시스템(색 선호)과 하향식 주의 시스템(글자 선택)의 경쟁이라고 설명하는데,12 실제로 이 이론은 stroop task를 할때 뇌가 어떤 활동 패턴을 보일지 예측했다.13 한편 이 이론은 주의의 본질적 특성상 한 감각 채널에서 주의가 가해지면 이게 다른 감각 채널에서도 주의를 부를 수 있다고 예측하는데, 왜냐하면 이미 두드러진 신경 활동은 다른 채널에도 간섭할 수 있기 때문이다. 이는 실제로 뇌파 연구에서 나타났다.14
주의 이론
선택적 주의가 작동하는 방식에 대한 이론은 3가지가 있다. 모두 경험적 증거가 있으며, 나중 이론이 제일 견고하다. 가장 먼저 나온 이론은 broadbent's filter theory(broadbent model)로, 이 이론은 선택적 주의가 정보입력 초기 단계에서 작동한다고 본다. broadbent's filter theory에서는 뇌에 입력된 정보가 다음의 과정을 거친다고 제안한다.
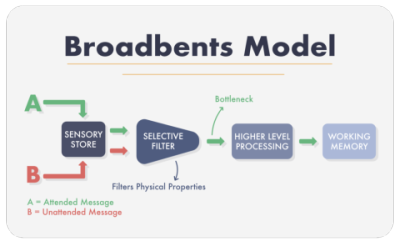
이론에 따르면, 외부에서 들어온 정보들은 일단 감각기억으로 저장된다. 그리고 이들 정보들은 단기기억으로 이동하는데, 도중에 filter와 detector를 거친다. filter에서는 들어온 정보를 목소리톤이나 성별 등을 통해 누가 말한 정보인지, 중요한 정보인지 판별한다. 이렇게 해서 중요하다고 판별된 정보(attended message)는 detector로 이동하고 그렇지 않은 정보(unattended message)는 차단된다. 그러면 detector에서 들어온 정보를 고차적으로 처리하여 단기기억으로 보낸다.
broadbent model은 여러가지를 설명하나, 칵테일 파티 효과(cocktail party effect)15를 잘 설명하지 못한다. 칵테일 파티 효과는 혼잡한 파티장에서 누군가 내 이름을 부르는 것을 바로 알아듣는 현상인데, 특별히 주의를 기울이지 않는 상황에서도 복잡한 소음 속에서 이름을 간파하기 때문에 특이한 현상으로 여겨졌다. 특히 이름은 성별도 목소리톤도 아니기 때문에 broadbent model과는 특히 안맞는다. 비슷하게 다른 실험16에서 연구자들은 피험자에게 어떤 문장을 양쪽 귀에 분산해서 들려주었다. 즉 '나는 개가 좋다'라는 문장을 말하는 경우 왼쪽 귀에는 '나는'과 '좋다'가 들리고, 그 사이에 오른쪽 귀에서 '개가'가 들리는 식이다. 목소리톤과 성별과는 관련이 없기 때문에 피험자는 한쪽 귀에서 나온 단어(와 혼합시킨 숫자들)만 들어야 하지만, 그들은 완전한 문장을 기억했다.
attenuation theory(intermediate selection model)은 broadbent model이 몰락한 후 나온 이론이다. 이는 유명한 인지과학자인 triesman이 제안했는데, 이 이론에서는 중요한 정보와 아닌 정보가 초기부터 분리된다는 점은 broadbent model과 비슷하지만 정보처리의 후기단계에서도 분리가 나타날 수 있다고 제안하였다. 이론에서 제안한 순서도는 다음과 같다.
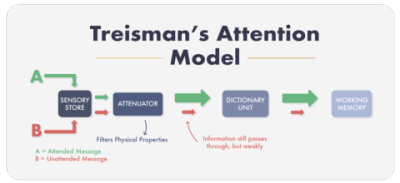
기본적인 구조는 broadbent model과 유사하다. 그러나 attenuation theory에서는 attenuator라는 시스템이 정보를 선별하는데, attenuator는 중요하지 않은 정보를 차단하는 대신 중요한 정보를 강화하고 아닌 정보를 약화한다. 또한 attenuator는 소리의 물리적 특성은 물론 음운적 특성과 의미도 중요한 기준으로 삼는다. 이렇게 강화되고 약화된 정보들은 모두 dictionary unit으로 가는데, 정보 차단은 여기서 이뤄진다. dictionary unit은 특정 역치에 다다른 단어는 단기기억으로 보내지만, 그렇지 못한 단어는 차단한다. 이 과정에서 중요하지 않은 정보가 걸러지지만, 만약 흔히 듣거나 중요한 정보와 관련된 단어들(이름은 여기에 해당한다)은 상위 기능에 의해 역치가 낮게 설정되어 통과가 가능하다.
마지막으로 late selection model은 1973년 Mackay가 제안한 이론으로, 위와 같이 선행적이거나 후행적인 선택기제를 가정하지 않는 이론이다. 이 이론을 제안한 Mackay는 피험자들에게 'They were throwing stones at the bank.'란 문장을 들려주었는데, bank는 은행과 둑의 의미를 같이 가지고 있다. 피험자들은 이 문장에 주의를 기울이는 한편, 다른쪽 귀에서는 river나 money가 들렸다. 실험 결과 피험자들은 자신이 river를 들었는지 money를 들었는지에 따라 bank를 다르게 해석했으며, 놀랍게도 다른쪽 귀에서 무슨 단어가 들렸었는지는 기억하지 못했다. 이는 걸러지는 정보와 아닌 정보의 경계가 모호할 수 있음을 보여준다.
late selection model은 load theory of attention17으로 체계화되었다. load theory에서는 정보가 선별되는 복잡한 경로를 가정하는 대신, 인간의 전체 주의력이 제한된 용량을 가지고 있다고 제안한다.(processing capacity) 이러한 관점에서 보면 주의가 필요한 과제들은 주의력이 많이 필요하거나(high-load) 적게 필요한(low-load) 과제로 나눌 수 있는데, low load한 과제를 수행하는 경우 주의력 자원이 남기 때문에 충분히 다른 정보(선택적 주의를 기울이지 않은)도 들을 수 있다. 하지만 매우 어려운 과제(high load)를 수행하는 경우에는 남은 주의력 따위는 없기 때문에 divided attention이 불가능하다. 그러한 경우 주의가 기울여지지 않은 자극은 모든 억제하여 주의력을 아끼는데, 실제로 움직이는 점(방해 자극)이 있었던 연구18에서는 주의력이 많이 필요한 상황에서는 움직임 지각과 관련된 MT의 활동이 감소하였다. 다른 여러 연구들19도 일관되게 주의에서 주의력이 소모되는 정도(load)가 주의와 깊게 관련되어 있다는 점을 지지했다.
주의의 발생과정
주의(선택적 주의)는 외부에서 자극이 입력될 때 발생한다. 외부에서 자극이 들어오면 먼저 들어온 자극이 주의를 일으킬수도 있고(early-selection viewpoint),20 이후 자극이 완전히 처리된 이후 다시 한번 주의가 일어날 수도 있다.(late-selection viewpoint)21 이 두 관점은 원래 대립했지만, ERP 연구22는 둘 다 맞다는 것을 보여주었다. ERP 연구는 같은 자극을 연달아 제시하고 이에 반응하는 뇌파를 측정하였는데, 주의가 집중되었음을 의미하는 P50은 보통 자극 출현 이후 80-100ms에 일어나지만 두번째 자극의 경우에는 35-85ms 후에 일어나 반응 시간이 더 짧아졌다. 이는 뇌가 정보처리가 완료되기 이전에 자극에 주의를 기울였다는 뜻으로, early-selection viewpoint를 지지한다고 할 수 있다.
다른 ERP 연구에서는 주의 집중과 관련된 뇌파를 조사했는데, 이 연구에서 피험자는 특정 톤(target tone)을 듣고 그 수를 세야 했는데, 이때 한쪽 귀(주로 왼쪽)에만 소리가 나타나니 한쪽 귀에만 주의를 기울이라는 지시를 받았다. 그리고 실제로는 다른쪽 귀에도 소리를 들려주고, 소리가 들려온 귀에 따라 뇌파가 달라지는지 관찰하였다. 실험 중 피험자는 한쪽 귀에만 주의를 기울였으니, 이러한 실험은 사람들이 선택적 주의를 기울이는 경우만 관련된 뇌파를 찾아내는데 수월할 것이다. 실험결과23 선택적 주의가 기울여진 경우 뇌파는 80ms 후 더 큰 음전하를 보였으며, 이 신호는 Nd(Negative difference)로 명명되었다. Nd는 청각자극을 활용한 연구뿐만 아니라 다른 자극을 사용한 연구24에서도 주의를 기울일때 나타났다.
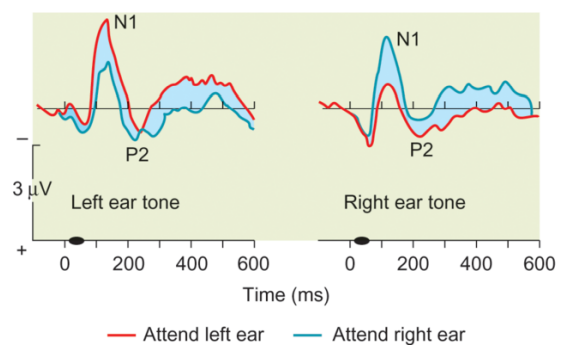
Nd가 early-selection viewpoint와 관련되었다면, N2pc는 late-selection viewpoint와 관련되어 있다. 음전하 방향의 요소인 N2pc는 자극 출현 이후 180-280ms 이후에 나타나는데, 자극 정보가 지각적 처리를 완벽히 거친 후 거기에 인위적으로 집중해야 할 때 주로 발생한다.25 비슷하게 P300은 사람들이 주의를 기울이고 대상을 모니터링할때 나타나는데,26 N2pc와 비슷하게 무언가 일을 하기 위해 대상에 주의를 기울여야 할 때(즉 작업기억을 사용할 때) 나타난다.27
선택적 주의(selective attention)
선택적 주의(selective attention)는 목표하는 특정 대상에만 선택적으로 기울여지는 주의를 말한다. 인간이 환경을 지각하는 경우에는 선택적 주의를 기울이는 대상에 지속적으로 초점이 고정되기 때문에 아이트래커를 사용하여 선택적 주의가 기울여지는 대상을 알 수 있다. 선택적 주의는 그 근원에 따라 bottom-up과 top-down으로 나눌 수 있는데, bottom-up attentional selection은 지각된 환경 자극이 너무 독특하거나 우리의 본성에 부합해서 우리의 주의를 끌어오는 경우이고, top-down attentional selection은 의도적으로 특정 대상에 선택적 주의를 기울이는 경우를 말한다.
주의의 전환28
주의를 전환하는 것은 한 대상에 기울이고 있던 주의를 철회하고, 다른 대상에 새로 주의를 기울이는 2단계의 과정으로 일어난다. 이는 fixation을 통해 관찰할 수 있는데, 왜냐하면 fixation이 가해지는 곳에 주의가 가해지기 때문이다. fixation이란 saccade의 일부로, saccade란 안구운동(eye movement)의 일종으로 급작스럽게 초점이 움직이는 안구운동을 saccade라 한다. 주로 아이트래커로 측정되는 안구운동은 무작위로 급작스럽게 초점이 움직이는 saccade 기간과 특정 지점에 초점이 멈춰있는 fixation 기간으로 나뉘는데, 이때 주로 fixation 기간에 초점이 향하는 곳에 대개 주의가 집중되며 이러한 주의를 overt attention이라 한다.
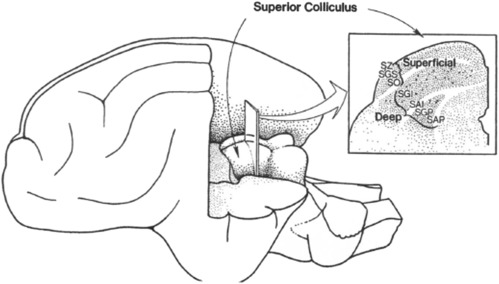
주의의 전환을 담당하는 뇌 영역은 superior colliculus이다. 실제로 superior colliculus와 basal ganglia가 손상된 supranuclear palsy 환자들은 의사의 지시에 따라 무언가를 보는 것은 잘해도 스스로 무언가를 보는 것에는 어려움을 느끼며,29 뇌영상 연구에서도 주의를 잘 끌어들이는 자극을 볼 때 superior colliculus 활동이 커졌다.30 superior colliculus는 saccade를 통제하여 주의를 전환하고는데, saccade란 안구운동(eye movement)의 일종으로 급작스럽게 초점이 움직이는 안구운동을 saccade라 한다. 주로 아이트래커로 측정되는 안구운동은 무작위로 급작스럽게 초점이 움직이는 saccade 기간과 특정 지점에 초점이 멈춰있는 fixation 기간으로 나뉘는데, 이때 주로 fixation 기간에 초점이 향하는 곳에 대개 주의가 집중되며 이러한 주의를 overt attention이라 한다. 물론 superior colliculus는 시각뿐만 아니라 다른 감각과 관련해서도 주의를 전환하는 것과 관련되어 있다.31
saccade는 express saccade와 regular saccade로 나눌 수 있다. express saccade는 주로 외부자극을 지각했을때 120ms 후에 나타나는 운동으로, 상향식 주의와 관련되어 있으며 superior colliculus에서 담당한다. 반면 regular saccade는 외부자극을 지각했거나 특정 대상에 주의를 기울일 때 나타나는데, 주로 주의를 기울이려고 하고 200-300ms 후에 나타나며32 FEF(Frontal Eye field)33에서 담당한다.
superior colliculus는 그 세부 부위에 따라 담당하는 기능이 약간씩 다르다. 가량 superior colliculus의 가장 바깥층(superficial layer)은 시각 자극의 출현과 사라짐, 움직임에 빠르게 반응하고, 반대쪽 신체의 retinotopic map이 표상되어 있다. 그렇기 때문에 superficial layer는 외부의 시각 자극을 찾고 위치를 식별하는데 용이하다.34 반면 다른 층은 intermediate layer와 deep layer가 있는데, 이 층은 대상의 감각적 특성에 민감하고 그곳을 향해 눈을 움직이게 한다. 실제로 이 층은 안구운동의 통제와 밀접하게 연관된 뇌간 영역과 연결되어 있다.35 즉 요약하면 superior colliculus 안에서도 superficial layer는 시각자극을 탐지하는 것과 관련되었고, 반면 intermediate layer와 deep layer는 그것을 향하도록 눈을 통제하는 것과 관련되어 있다.
한편 주의 전환은 superior colliculus뿐만 아니라 SPL(Superior Parietal Lobe)에서도 담당한다.36 superior colliculus와 마찬가지로 SPL도 주의를 전환하는 것과 관련되는데, 특정 감각 영역에서의 주의 전환 뿐만 아니라 서로 겹쳐진 그림처럼 지각적 차원에서 서로 다른 대상에 주의를 기울여야 할 때도 관여한다.37 연구에 따르면 기존 대상에서 주의를 철회하고 다른 대상에 주의를 주는 두 과정은 SPL의 서로 다른 영역에서 일어난다.38
overt attention과 covert attention
overt attention은 일종의 내적 주의로 번역되는데, 개인이 스스로 기울이는 선택적 주의를 말한다. 사실 많은 경우 사람들은 자신이 보고자 하는 무언가에 주의를 기울인다. 이러한 주의는 모두 overt attention이라 할 수 있다. 정확히 학자들은 안구운동을 통해 측정되는 주의를 overt attention이라고 여기는데, 안구운동 중 fixation 기간에 초점이 집중되는 곳이 overt attention이 가해진다고 여긴다. 사실 이렇게 주의가 가해지는 곳은 주의가 직접 가해지기 이전부터 fixation이 시작된다.
overt attention은 top-down processing의 결과물로, 상위 인지기능에 의해 유발된 주의이다. 비슷한 경우가 scene schema인데, scene schema는 시야에 들어온 장면에 대한 지식(전형적인 면에 대한 지식)을 말한다. 이는 보통 순서의 형태로 저장되어 있는데, fixation도 이 순서에 맞게 가해진다. 가령 비밀번호가 1958인 자택 현관문을 열고 있다면, fixation이 1번->9번->5번->8번 순으로 진행될 것이다. 그래서 이론상 아이트래커만 있으면 타인의 비밀번호를 알아낼 수 있는데, 물론 모르는 사람이 나에게 아이트래커를 씌우려 한다면 보통 경찰에 신고할 것이다. 한편 top-down processing이 주의를 유발한다면 bottom-up processing도 주의를 유발할 것이다. 어떤 자극들(특히 색이나 움직임)은 주의를 일으키고 자신들에게 끌어모은다.
그러나 어떤 주의는 안구운동으로 측정되지 않으며, 이들을 covert attention이라 하나 이러한 구분은 개론에나 나오지 실제 연구현장에서는 잘 쓰이지 않는다. overt하지 않은 주의는 precueing이 있는데, precueing은 피험자가 무언가 나타나리라고 기대하는 곳에 초점이 가해지지 않아도 주의가 가해지는 현상을 말한다. 이를 측정하는 실험에서 연구자는 피험자에게 화살표 등을 통해 어떤 지점에 주의를 기울이도록 유도하는데, 실제 측정을 해보면 그곳에 초점이 가해지지 않아도 피험자들은 그 지점에서 출몰하는 자극을 더 잘 지각한다.
멀티태스킹(divided attention)
멀티태스킹(multi-tasking, 다중작업)은 여러가지 인지적 작업을 동시에 처리하는 것을 의미한다. 멀티태스킹은 과제를 매우 빠르게 직렬적으로 처리하거나, 여러 과제를 동시에 병렬처리하는 경우에 가능하다. 전자는 2020년 현재의 컴퓨터고, 후자는 생물체의 뇌이다. 인간은 오감을 통한 지각과 사회적/작업적 정보처리를 동시에 수행하며, 이는 인간의 뇌에 이들을 처리하는 별개의 회로가 동시에 작동하기 때문이다.
비록 자동적 정보처리는 대부분 병렬적으로 이뤄지지만, 의식적 정보처리는 직렬적으로 수행된다. 이는 사람이 실생활에서 멀티태스킹을 잘 못하는 이유이다. 실제로 사람들은 여러 인지과제에서 멀티태스킹을 힘들어하는 모습을 보여준다. 한 실험에서 피험자들은 운전중에 라디오를 듣거나 전화통화를 할때(자동차 오디오로 하더라도) 더 늦게 브레이크를 밟고 정지판을 더 늦게 보았으며,39 실제 운전 현장에 대한 연구에서도 비슷한 결론이 나왔다.40 다른 연구에서 피험자는 시각정보와 청각정보를 동시에 처리해야 했는데,41 fMRI 측정 결과 주의가 청각으로 기운 경우에는 시각피질이, 주의가 시각으로 기운 경우에는 청각피질이 활동이 감소하였다. 이는 두 과제에 동시에 주의를 잘 기울인다는 말이 거짓에 가까움을 보여준다. 어떤 연구에서는 피험자가 두 게임을 동시에 하는 경우 오류율이 하나만 할때보다 8배나 증가하였다.42 이는 자신이 멀티태스킹을 잘 한다고 주장하는 사람들의 주장과 상반되는데, 흥미롭게도 일상에서 멀티태스킹을 자주 한다고 보고한 사람이 정작 멀티태스킹 과제에서는 다른 사람보다 저조한 수행을 보였다.43
그러나 충분히 숙련되면 제한적으로나마 멀티태스킹이 가능할 수도 있다. 멀티태스킹에 대한 초기 연구44에서 연구자들은 피험자들에게 멀티태스킹 과제를 시켰다. 물론 예상했듯이 피험자들은 형편없이 못했고, 성공률은 50%에 근접했다.(우연 수준이 50%이다) 그러나 과제를 충분히 반복하자 나중에는 80%까지 성공률을 올리는데 성공했다. 이는 멀티태스킹도 부분적으로는 숙달이 가능하다는 점을 보여준다. 다만 2가지를 주의해야 하는데, 먼저 이 실험은 일상생활보다 간단한 실험실 과제를 사용했고, 숙달되기까지 1200번을 해야 했다. 가능은 하지만, 쉬운 일은 아니다.
object-based visual attention45
안구운동 연구자들이 관찰하는 주의는 대개 location-based attention이다. 이 주의는 어떤 위치에서 다른 위치로, 또 다른 위치로 옮겨간다. 그러나 어떤 경우에는 주의가 가만히 고정되어 있기도 하다. 어떤 경우에는 초점은 고정되어 있지만, 주의에 들어오는 정보는 달라진다. 그중 초점은 고정되어 있으면서 초점을 둔 지점에 대해 점점 더 세세하고 물체와 관련된 정보에 주의가 기울어지는 경우 이를 object-based visual attention이라 한다.
egly와 동료들의 1994년 연구에서 연구자들은 두 막대를 제시하고 막대 끝에 빛이 들어오면 말하라고 지시하였다. 실험에서 사용된 막대는 다음과 같았다.
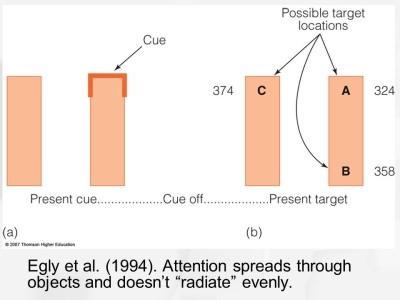
실험결과, A에서 빛이 나올때 가장 반응이 빨랐다. 이는 기존 이론에 부합하는 결과이다. 그러나 그 다음으로 B에서 빛이 나올때 반응이 빨랐다. A-B간 거리는 A-C간 거리와 같기 때문에 이는 location-based attention으로는 설명할 수 없으며, 이러한 결과는 A와 B가 있는 막대가 하나로 인식되어 막대 내부에서 object-based visual attention이 작동하기 쉬웠기 때문에 나타났다.
object-based visual attention은 배경이 어엄청 고요하거나, 몇개의 물체만 있는 경우 일어나기 쉽다. 또한 배경이 복잡하더라도, 물체가 매우 역동적으로 움직이는 경우에도 일어나기 쉽다.
인간이 기울이는 주의는 2가지를 중심으로 주어진다. 사람들은 특정 장소에 주의를 기울이거나(space-based viewpoint of attention), 특정 대상에 주의를 기울인다.(object-based viewpoint of attention)46 이 중 object-based visual attention은 후자가 두드러진 결과이다. object-based visual attention에 대한 다른 연구47에서 피험자는 서로 겹쳐있는 얼굴과 집 사진을 봤는데, 얼굴에 주의를 기울이는 경우 FFA가 활동했고 집을 보는 경우 parahippocampal place area가 활동했다. 이는 위 연구와 마찬가지로 물체와 같은 특정 대상이 장소와 마찬가지로 주의를 기울이는 대상이 될 수 있음을 보여준다.
space-based viewpoint of attention은 주로 extrastriate cortex의 시각 중추(V1-V4)에서 연산되며,48 특히 시각 중추에 있는 retinotopic map을 토대로 위치를 계산한다. 이러한 처리는 P1 신호와 관련되어 있는데, P1 신호는 자극을 지각한 이후100ms에 출현하며 이미 주의를 기울이던 장소에 자극이 출현하면 더 강해진다.49
object-based viewpoint of attention은 색과 같은 물체의 특성을 중심으로 일어나는데, 색의 경우 색지각을 연산하는 V4의 활동이 색을 중심으로 한 주의 기울이기와 관련되어 있다.50 모양의 경우는 물체의 모양을 처리하는 시각의 ventral stream 쪽 영역이 활동했고 속도에 주의를 기울이는 경우 속도 관련 정보를 처리하는 MT에서 활동했는데, 즉 space-based viewpoint of attention의 경우와 마찬가지로 특정 장소나 대상에 주의를 기울이기 위해서는 해당 장소나 대상을 지각하는 영역의 정보처리가 바탕이 되어야 한다.
한편 과거 연구51에서는 object-based viewpoint of attention이 자극 출현 이후 200-300ms에 나타나 space-based viewpoint of attention보다 느리다고 여겨졌지만, 최근 연구52에서는 색을 중심으로 지각하는 경우 방해 자극(색)이 많은 경우에 P1이 더 크게 나타나 object-based attention도 space-based attention만큼 일찍 나타날 수도 있다고 주장하였다. 다른 연구(n=11)53에서도 피험자가 얼굴에 주의를 기울일때 발생하는 M170이 space-based attention 때의 P1만큼 빠르게 나타났다고 보고했다.
생리적 측면54
다른 기능이 그러하듯이, 시상은 주의에서도 외부자극을 제련하여 피질로 보내는 역할(gating, gate-keeping)을 한다. 이러한 역할은 시상에서도 LGN과 pulvinar가 담당하는데,55 가령 LGN은 피질의 명령을 반영하여 시각적 자극에 선택적 주의를 기울이는 역할을 한다.56 pulvinar는 선택적 주의와 관련된 여러 영역들을 연결하는 것으로 보이는데, 실제로 원숭이 연구에서 pulvinar는 시각 자극의 탐지 및 주의 배분과 관련된 V4 및 inferotemporal cortex와 동시에 활동하였다.57 이러한 synchrony는 알파파 대역인 8-15Hz에서 잘 드러났다. 또한 pulvinar는 방해되는 자극을 무시하고 주의를 목표하는 대상에만 기울이도록 만드는데,58 그래서 pulvinar가 손상되면 방해 자극 속에서 목표 자극에 주의를 기울이는 것에 어려움을 느낀다.59
선택적 주의는 주로 두정엽에서 담당하지만, 이를 직접 행동으로 옮기는 것은 전두엽이다. 특히 전두엽의 SMA(supplementary Motor Area)와 Anterior Cingulate가 이 기능을 담당한다. 그래서 전두엽이 손상된 환자들 중 일부는 motor neglect를 보이기도 하는데, motor neglect는 hemineglect의 일종으로 한쪽 측면의 자극에 반응하여 행동을 보이지 못하는 증상을 말한다.60 motor neglect 환자는 별다른 motor paralysis를 겪지 않으며, 지각에 있어서는 hemineglect를 보이지 않는다.61 또한 연구들은 원숭이의 cingulate motor area의 rostral한 영역에 위치한 뉴런이 원숭이가 운동 단서를 포착했을때 활동했고,62 인간 뇌전증 환자를 대상으로 한 EEG 연구63에서도 환자가 주의를 준 자극에 반응하기 직전과 직후에 SMA 일부와 cingulate의 dorsal한 영역에서 베타파가 관측되었다.
또한 regular saccade와도 관련된 FEF는 하향식 주의를 일으키는 것과도 관련되어 있다.64 이와 관련된 연구65에서 FEF는 주의를 기울이기 전에도 두정엽의 SPL과 함께 주의 관련 단서가 나타났을때 강하게 활동하였다. 이는 FEF가 후술할 SPL과 마찬가지로, 선택적 주의를 조작하고 일으키는 영역일 수 있음을 보여준다. 이에 대한 다른 연구66에서 연구자는 마카크 원숭이에게 색을 구분하는 과제를 시켰는데, 과제를 위해 주의를 기울이는 동안 FEF의 뉴런은 V4의 뉴런과 함께 같은 감마파를 보였다. 이 신호는 FEF가 8-13ms 더 빨랐고, FEF를 손상시키자 동기화와 주의를 기울이는 능력이 모두 감소하였다. 인간연구67에서도 마찬가지로 얼굴에 주의를 기울이는 사람은 PFC와 FFA가 동기화되었고, 집에 주의를 기울이는 사람은 PFC와 PPA가 동기화되었으며, 두 경우 모두 PFC의 신호가 20ms 더 빨랐다.
ACC는 주로 자동적 정보처리 시스템 간에 충돌이 발생하거나 일을 수행하는데 있어 오류가 발생했을때 활동하는데, 집행기능의 일부이기 때문에 주의와도 관련되어 있다. 선택적 주의에서 ACC는 선택적 주의를 일으키는 일을 하는데, ACC가 활동할 만한 문제상황이 발생했을때 선택적 주의를 해당 문제에 집중시키는 것이 ACC의 일이다. ACC뿐만 아니라 supplementary eye field도 여러 자동적 정보처리 시스템이 충돌하는 상황(response conflict)에 활동한다.68
한편 DPC와 VPC는 두정엽에 위치하지만, 두정엽에만 있진 않는다. 이들은 대뇌 곳곳에 가지를 뻗어 자신들만의 네트워크를 형성한다. 주의의 분배와 정보처리는 이들 네트워크를 통해 이루어진다. fMRI 연구에 따르면 VPC는 STG와 temporal parietal junction, 그리고 VLPFC, DLPFC와 함께 네트워크를 형성하는데 반해 DPC는 FEF(Frontal Eye Field), IPS와 함께 네트워크를 형성한다. 신기한 사실은 하향식 처리를 명령하는 DPC가 정작 연결된 부위는 실제 하향식 처리를 담당하는 PFC가 아니라 FEF같은 superior frontal cortex이다.
두정엽의 역할
기본적으로 주의의 배분은 두정엽이 담당하며, 특히 우측 두정엽이 담당한다. 두정엽은 주의를 적절한 곳에 할당하고 유지(sustained attention)하는데, 각자 다른 영역이 주의 배분에 관여한다. 가령 주의가 가해지는 시간과 공간에 따라 활동하는 영역도 달라지지만 거의 모든 종류의 주의 배분에는 항상 right IPS와 inferior parietal cortex가 활동한다.69 그리고 TPJ(TemporoParietal Junction)도 주의 배분에 관여하는데, 그래서 TPJ가 손상되면 P300 신호가 일어나지 않는다.70
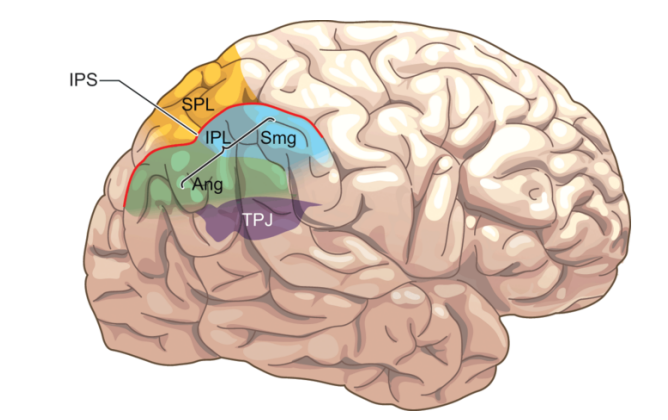
두정엽은 주의를 배분하기 위해 pulvinar에서 올라온 정보를 다시 한번 처리한다. 이러한 처리는 SPL(Superior Parietal Lobule)과 IPL(Inferior Parietal Lobule), intraparietal region에서 담당하는데, SPL은 주로 하향식 처리를 맡고,71 IPL은 상향식 처리를 맡는다.72 그래서 SPL은 주의를 집중하고 전환하는 선택적 주의를 담당하고, 반면 IPL은 주의를 끄는 지각적 대상에 주의를 끌리게 만든다. 하향식 주의는 주로 Dorsal Parietal Cortex(DPC)에서 처리하고 상향식 주의는 Ventral Parietal Cortex(VPC)에서 처리하는데, 외부 자극이 주의를 끌때는 VPC가 활동하는 반면 집행 기능을 이용한 하향식 처리가 필요할때는 DPC가 활동한다. 그리고 이 둘은 intraparietal region에서 만나 서로의 기능을 조절한다.
SPL와 IPL은 각각 하향식 주의와 상향식 주의의 핵심이기 때문에, SPL은 주의의 분배에 관여하는 반면 IPL은 외부에서 나타난 대상에 주의를 기울이도록 만드는게 핵심 기능이다. 가령 IPL은 사람이 미처 인식하지 못했던 자극을 인식했을때 활동하는데,73 그래서 예상치 못한 장소에 나타난 자극이나 매우 드물게 발생하는 사건을 지각했을때 활성화된다.74 실제로 이 주변이 손상된 사람들은 hemineglect를 앓는데,75 hemineglect란 사람의 한쪽 측면(왼쪽이나 오른쪽)에 주의를 기울이지 못해 그 쪽 측면의 모든 자극을 무시하는 증상이다. IPL 주변이 손상된 hemineglect 환자들은 의사가 직접 특정한 무언가에 주의를 기울이라고 지시하면 대상을 인식할 수 있지만, 그러한 (하향적)단서가 주어지지 않는 경우 무슨 일이 일어나도 무시되는 측면의 대상을 지각하지 못한다.76
앞서 말했다시피 intraparietal region(intraparietal sulcus와 intraparietal lobe)은 SPL와 IPL의 정보를 통합하는데, 이 영역은 두 영역의 정보를 통합해서 어느 곳에 먼저 주의를 기울여야 할지를 보여주는 일종의 priority map을 만드는 것으로 보인다.77 그래서 intraparietal region은 하향식 주의나 상향식 주의가 나타나는 모든 경우에 활동하며,78 궁극적으로 priority map에 기초하여 사람이 시각 자극에 선택적 주의를 기울이게 만든다.79 이 priority map은 주의가 기울여질 공간에 대한 완전판 정보라고도 할 수 있는데, 이 지도상에서는 색이나 선과 같이 주의를 끄는 자극도 크게 나타나고, 선택적 주의가 기울여진 자극도 크게 나타난다. 한편 intraparietal region은 FEF(Fronto Eye Field)와 같은 운동 관련 영역과도 긴밀히 연결되어, 주의가 기울여진 곳에 적절한 운동이 잘 통합되도록 작동한다.
변화맹(change blindness, inattentional blindness)
변화맹(change blindness)은 주의가 가지는 역할을 잘 보여주는 예시로, 사람들이 세부적인 변화를 잘 못보는 현상이다.80 이에 대한 고전적인 연구들이 꽤 있지만, 많은 연구자들은 세부적인 차이만 있는 두 사진을 0.05초 간격으로 보여주고 변화가 있는지 물어보는 것으로 변화맹 여부를 확인한다.
이를 보여주는 대표적인 실험이 고릴라 실험81이다. 이 실험에서 피험자들은 사람들이 농구를 하고 있는 영상을 보고 그들이 패스를 총 몇 번 하는지 세어달라고 지시받았다. 영상이 끝나자 피험자들은 꽤 정확하게 세었는데, 이때 연구자가 피험자에게 영상에서 중간에 지나간 고릴라를 봤냐고 질문했다. 실제로 영상 중간에 고릴라 옷을 입은 사람이 나와 중앙에서 가슴을 두드리고 지나갔지만, 피험자 대부분은 고릴라를 보지 못했다. 이는 피험자들이 패스 횟수에만 주의를 기울였기 때문에 고릴라가 주의에 포착되지 못해 지각되지 않았기 때문이다.
다른 실험82에서 실험자는 대학 캠퍼스에 있는 아무나에게 길을 물었다. 그리고 아무나가 질문에 답하는 동안 문을 든 두명의 인부(로 위장한 실험자)가 사이로 지나갔다. 인부가 지나간 후 피험자는 대답을 마쳤으며, 길을 묻고 대답하는 과정에서 어떤 이상도 느끼지 못했다. 반전은, 길을 묻는 사람이 바뀌었다는 것이다. 인부들이 문을 들고 사이를 지나갈 때, 질문을 던진 실험자는 문과 함께 사라지고 다른 실험자가 대신 거기 서있었다. 이런 뻔뻔한 속임수는 15명 중 8명이 알아차리지 못했다. 이런 현상은 실제 농구시합을 하는 상황에서도 발생할 수 있다.83
변화맹은 주의가 기울여지지 않아서 발생한다.84 실제로 변화맹은 사람이 변화한 대상에 주의를 잘 기울이지 못할 때 주로 나타나며, 주의를 잘 끄는 대상에서는 변화맹이 덜 일어난다.85 이는 무주의맹과도 비슷한데, 무주의맹(inattention blindness)은 주의가 기울여지지 않은 물체를 지각하지 못하는 현상이다.86 이 두 현상 모두 주의가 지각에서 얼마나 중요한지 잘 보여준다.
결합 문제(binding problem, feature integration theory, FIT)
결합 문제(binding problem)는 지각된 서로 다른 자극들이 왜 하나의 단일한 상으로 구성되는지에 대한 문제이다.87 인간은 물체의 색, 방향, 질감 등의 요소를 서로 떨어진 뉴런을 통해 지각한 후 이를 합친다. 문제는 이러한 정보를 어떻게 합쳐서 우리가 보는 완전한 상이 되는가에 대한 문제이다. 진화적 필요성으로 인해 그러한 기제가 진화한 것은 당연한 일이지만, 그것이 어떻게 되어있는가는 많은 학자들의 관심거리이다.
결합 문제는 착각적 접합(illusory conjunction)을 통해 주로 연구되었다. 착각적 접합은 서로 다른 시각 자극들이 잘못 결합되는 현상으로, 결합이 제대로 되지 못한 사례이다. 이에 대한 선구적인 연구88에서 연구자들은 검정 숫자들이 서로 다른 유채색 문자들을 둘러싸고 있는 시각자극을 피험자에게 매우 짧은 시간 보여주고, 그들에게 검정 숫자를 보고한 다음 유채색 숫자를 보고하도록 지시했다. 자극은 0.1초동안 나타났고 잔상이 남지 않도록 설계되었다. 실험 결과 피험자들은 유채색 문자들의 색을 헷갈려 했는데, 예를 들어 빨간 A와 파란 B가 제시되면 빨간 B와 파란 A로 착각하는 경우가 많았다. 이러한 경향은 일관되게 나타났으며 다른 오류보다 더 자주 나타났다. 같은 실험에서 문자들을 크기와 색, 모양으로 차별화한 도형을 바꾼 경우에도 결과는 동일했다.
트리즈먼과 동료들은 특징통합이론(feature integration theory ,FIT)으로 이를 설명하였다. 이 이론에 따르면 여러가지 자극이 통합되기 위해선 그러한 자극들에 주의가 가해져야 한다.89 즉 개개의 자극은 주의를 기울이지 않아도 충분히 인지되지만, 여기에 주의력이 배분되어야 자극의 연결이 가능하다는게 이론의 설명이다. 실제로 피험자들이 유채색 문자들에 특별히 주의를 기울이게 하자 착각적 접합은 사라졌다.90
FIT에 따르면 binding은 2단계를 거쳐 일어난다. 첫번째 단계는 preattentive stage인데, 이 단계에서는 개별 자극들이 각각의 감각경로를 통해 분석되고 지각된다. 이것은 무의식적으로 일어나고 주의가 필요하지 않다. 그 다음 단계가 초점 주의 단계(focused attention stage)인데, 이 단계에서 주의가 자극들에 가해지면서 binding이 일어난다. 위에 트리즈먼과 동료들의 실험에서도 실험하는 도형들에 주의를 집중하게 하자, 아주 짧은 시간에만 나타남에도 불구하고 착각적 접합은 나타나지 않았다. 비슷하게 balint's syndrome을 앓고 있어 주의집중이 힘든 환자 RM의 경우에도 착각적 접합을 매우 많이 일으켰다.
자극의 결합은 배측 경로91와 복측 경로92의 정보 모두에 의지한다. 이중에서도 후두엽-두정엽으로 이어지는 배측 경로가 결합 문제에서 중요한 일익을 담당하는 것처럼 보인다. 실제로 두정엽이 손상된 뇌손상 환자는 착각적 접합을 자주 일으켰으며,93 TMS를 통해 후두-두정 영역을 마비시키자 일반인도 착각적 접합을 자주 일으켰고 시각피질을 마비시켰을때는 그렇지 않았다.94 그리고 이렇게 두정엽이 손상된 사람들은 단일 자극(접합이 필요없는)은 잘 지각하여,95 이것이 자극 결합의 문제임을 보여주었다. 보다 세부적으로 자극의 결합은 두정엽의 intraparietal region에서 담당하는 것으로 보이는데, 이 영역은 하향식 주의와 상향식 주의를 통합하는 영역이기도 하다. 이에 관한 원숭이 연구96에서 원숭이의 intraparietal region을 비활성화자 원숭이들이 착각 접합을 일으켰다.
이 분야의 주요 연구자로는 로버트슨(robertson), 트리즈먼(treisman)이 있다.
주의의 생리적 기제97
각성은 대개 RAS(Reticular Activating System)에서 관장하는데, RAS는 뇌간에 위치한 여러 영역의 연결망으로 수면주기나 스트레스와 같은 각성과 관련된 다양한 기제를 관장하며 이 영역이 손상되면 코마가 일어난다. 코마는 주로 RAS가 손상되거나 기능이 저하될 때 나타나는데, 뇌수막염(meningitis)이나 대사장애, 가스중독, 티아민 부족, 독극물 중독 등에 의해 이러한 일이 일어날 수 있다. 이러한 요인들은 대개 뇌에만 영향을 끼치며, 그럼에도 불구하고 몸 전체의 기능을 저하시켜 아주 간단한 반사 작용들도 불가하게 만든다.
RAS는 서로 분리된 2가지 회로를 통해 신경전달물질을 대뇌에 전파해서 각성을 유발한다.98 한 회로는 dorsal route로, dorsal route는 tegmental nuclei에서 발원하여 thalamus의 medial dorsal/intralaminar/reticular nuclei를 거친 다음, 대뇌피질에 아세틸콜린을 전달하여 글루탐산을 활성화함으로서 대뇌피질의 각성을 유발한다. ventral route는 basal forebrain을 거쳐 대뇌피질에 각성을 유발하는데, 다시 서로 다른 2가지 회로로 나뉜다. 한 회로는 raphe nuclei에서 발원하여 세로토닌을 전달하고, 다른 회로는 locus coeruleus99에서 발원하여 노르에피네프린을 전달한다.
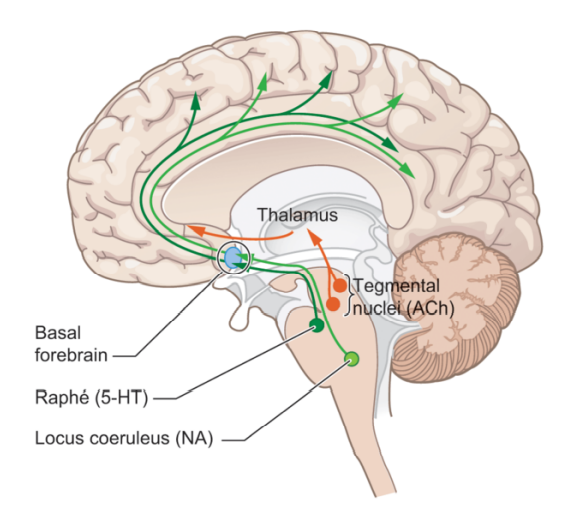
두 회로 중 각성에 가장 필수적인 역할을 담당하는 회로는 dorsal route이다. 실제로 코마 상태나 다른 최소의식 상태(minimally conscious state)는 dorsal route의 intralaminar nuclei나 다른 paralaminar 영역이 손상되었을때 발생하며 이 영역을 자극하면 원숭이의 각성이 증가한다.100 그러나 다른 회로도 각성에 일정 수준 기여하는데, 가령 locus coeruleus는 수면과 관련되어 있으며, single cell recording 연구에 따르면 locus coeruleus는 각성을 유발하는 자극이 주어졌을때 빠르게 활동했다. 이 영역의 손상은 높은 각성 수준에서의 정보처리를 어렵게 할 수 있다. 또한 노르에피네프린 회로는 스트레스 관련 활동과 관련되어 있고, 세로토닌 회로는 렘수면과 관련되어 있다.
vigilance는 위에서 나왔던 노르에피네프린 회로와 dorsal route가 관여된다. 각성과 마찬가지로 dorsal route는 vigilance에 중추적인 역할을 담당하며,101 노르에피네프린 회로는 주의를 끄는 자극에 반응하는 능력과 관련되어 있는데102 여기서 노르에피네프린 회로는 basal forebrain에 머물지 않고 다시 시상의 midline nuclei로 연결되어 vigilance에 영향을 끼친다. 이에 대한 한 연구103에서 midline nuclei의 활동 저하는 60-minutes auditory vigilance task에서의 수행저하와 연관되었다.
이러한 점들을 볼 때 시상은 각성과 주의를 연결하는 중계기 역할을 하는 것으로 보인다. 이에 대한 뇌영상 연구104에서 피험자들은 4분할된 화면에서 랜덤하게 등장하는 7에 주의를 기울여야 했는데, 피험자들은 각성 수준에 따라 3가지 그룹을 나뉘었다. 뇌영상 촬영 결과 각성 수준과 상관없이 피질 부위는 항상 활동한 반면, 시상의 ventrolateral 영역은 각성이 낮을때 크게 활동하고 각성이 높을때는 작게 활동했다. 이는 과제에 필요한 vigilance를 보충하기 위해 시상이 각성이 적은 상황에서는 vigilance를 일으켜 보충하고, 높은 상황에서는 쉬었기 때문에 나타난 결과로 보인다.
물론 위에서 보았듯이, 피질도 각성과 vigilance에 관여하며 특히 우뇌가 그러한 것으로 보인다. 실제 뇌손상 연구105에서 우뇌가 손상된 환자들은 과제가 청각적이건 시각적이건 주의 관련 수행이 더 떨어졌으며, 위험자극에 대한 심박수 변화도 느려졌다.106 10-15명을 대상으로 한 다른 연구107에서는 우뇌의 전두엽과 inferior parietal region에 자극을 주었는데, 이 연구도 우뇌가 vigilance에 중요한 역할을 한다고 보고하였다. 이러한 결과는 전기생리적 측정108에서도 재현되었기 때문에, 우뇌도 각성 및 vigilance에 중요한 역할을 하는 것으로 보인다.
- Banich, M. T., & Compton, R. J. (2018). Cognitive neuroscience. Cambridge University Press [본문으로]
- Reddy, L., Kanwisher, N. G., & VanRullen, R. (2009). Attention and biased competition in multi-voxel object representations. Proceedings of the National Academy of Sciences, 106(50), 21447-21452 [본문으로]
- Chun, M. M., Golomb, J. D., & Turk-Browne, N. B. (2011). A taxonomy of external and internal attention. Annual review of psychology, 62(1), 73-101. [본문으로]
- Li, F. F., VanRullen, R., Koch, C., & Perona, P. (2002). Rapid natural scene categorization in the near absence of attention. Proceedings of the National Academy of Sciences, 99(14), 9596-9601 [본문으로]
- Simons, D. J., & Chabris, C. F. (1999). Gorillas in our midst: Sustained inattentional blindness for dynamic events. perception, 28(9), 1059-1074 [본문으로]
- Wickens, C. D. (1980). The structure of attentional resources. Attention and performance VIII, 8, 239-257. [본문으로]
- Desimone, R., & Duncan, J. (1995). Neural mechanisms of selective visual attention. Annual review of neuroscience, 18(1), 193-222 [본문으로]
- Reynolds, J. H., Chelazzi, L., & Desimone, R. (1999). Competitive mechanisms subserve attention in macaque areas V2 and V4. Journal of Neuroscience, 19(5), 1736-1753 [본문으로]
- Pessoa, L., Kastner, S., & Ungerleider, L. G. (2003). Neuroimaging studies of attention: from modulation of sensory processing to top-down control. Journal of Neuroscience, 23(10), 3990-3998;Kastner, S., De Weerd, P., Desimone, R., & Ungerleider, L. G. (1998). Mechanisms of directed attention in the human extrastriate cortex as revealed by functional MRI. science, 282(5386), 108-111 [본문으로]
- Doesburg, S. M., Bedo, N., & Ward, L. M. (2016). Top-down alpha oscillatory network interactions during visuospatial attention orienting. Neuroimage, 132, 512-519 [본문으로]
- Mozer, M. C., & Sitton, M. (1998). Computational modeling of spatial attention. Attention, 9, 341-393 [본문으로]
- O'Reilly, R. C., Munakata, Y., Frank, M. J., & Hazy, T. E. (2012). Computational cognitive neuroscience (Vol. 1124). Mainz: PediaPress;Cohen, J. D., Braver, T. S., & O´ Reilly, R. (1996). A computational approach to prefrontal cortex, cognitive control and schizophrenia: recent developments and current challenges. Philosophical transactions of the royal society of london. Series B: Biological sciences, 351(1346), 1515-1527 [본문으로]
- Herd, S. A., Banich, M. T., & O'reilly, R. C. (2006). Neural mechanisms of cognitive control: An integrative model of Stroop task performance and fMRI data. Journal of cognitive neuroscience, 18(1), 22-32 [본문으로]
- Müsch, K., Hamamé, C. M., Perrone-Bertolotti, M., Minotti, L., Kahane, P., Engel, A. K., ... & Schneider, T. R. (2014). Selective attention modulates high-frequency activity in the face-processing network. Cortex, 60, 34-51 [본문으로]
- Schactor 외 2명,'심리학 입문(2판)',민경환 외 8명 역,시그마프레스,2015 [본문으로]
- Gray, J. A., & Wedderburn, A. A. I. (1960). Shorter articles and notes grouping strategies with simultaneous stimuli. Quarterly Journal of Experimental Psychology, 12(3), 180-184. [본문으로]
- Lavie, N., & Dalton, P. (2014). Load theory of attention and cognitive control. The Oxford handbook of attention, 56-75 [본문으로]
- Rees, G., Frith, C. D., & Lavie, N. (1997). Modulating irrelevant motion perception by varying attentional load in an unrelated task. Science, 278(5343), 1616-1619 [본문으로]
- Lavie, N., & Tsal, Y. (1994). Perceptual load as a major determinant of the locus of selection in visual attention. Perception & psychophysics, 56(2), 183-197;Lavie, N. (1995). Perceptual load as a necessary condition for selective attention. Journal of Experimental Psychology: Human perception and performance, 21(3), 451 [본문으로]
- Broadbent, D. E. (2013). Perception and communication. Elsevier. [본문으로]
- Deutsch, J. A., & Deutsch, D. (1963). Attention: Some theoretical considerations. Psychological review, 70(1), 80. [본문으로]
- Eimer, M. (2014). The neural basis of attentional control in visual search. Trends in cognitive sciences, 18(10), 526-535. [본문으로]
- Hillyard, S. A., Hink, R. F., Schwent, V. L., & Picton, T. W. (1973). Electrical signs of selective attention in the human brain. Science, 182(4108), 177-180. [본문으로]
- Desmedt, J. E., & Robertson, D. (1977). Differential enhancement of early and late components of the cerebral somatosensory evoked potentials during forced‐paced cognitive tasks in man. The Journal of physiology, 271(3), 761-782;Van Voorhis, S., & Hillyard, S. A. (1977). Visual evoked potentials and selective attention to points in space. Perception & psychophysics, 22(1), 54-6 [본문으로]
- Luck, S. J., & Hillyard, S. A. (1994). Spatial filtering during visual search: evidence from human electrophysiology. Journal of Experimental Psychology: Human Perception and Performance, 20(5), 1000 [본문으로]
- Donchin, E. (1981). Surprise!… surprise?. Psychophysiology, 18(5), 493-513 [본문으로]
- Wickens, C., Kramer, A., Vanasse, L., & Donchin, E. (1983). Performance of concurrent tasks: a psychophysiological analysis of the reciprocity of information-processing resources. Science, 221(4615), 1080-1082;Kramer, A. F., Wickens, C. D., & Donchin, E. (1985). Processing of stimulus properties: evidence for dual-task integrality. Journal of Experimental Psychology: Human Perception and Performance, 11(4), 393 [본문으로]
- Banich, M. T., & Compton, R. J. (2018). Cognitive neuroscience. Cambridge University Press [본문으로]
- RAFAL, R. D., POSNER, M. I., FRIEDMAN, J. H., INHOFF, A. W., & BERNSTEIN, E. (1988). Orienting of visual attention in progressive supranuclear palsy. Brain, 111(2), 267-280 [본문으로]
- Anderson, E. J., & Rees, G. (2011). Neural correlates of spatial orienting in the human superior colliculus. Journal of Neurophysiology, 106(5), 2273-2284 [본문으로]
- Fairhall, S. L., & Macaluso, E. (2009). Spatial attention can modulate audiovisual integration at multiple cortical and subcortical sites. European Journal of Neuroscience, 29(6), 1247-1257 [본문으로]
- Schiller, P. H., Sandell, J. H., & Maunsell, J. H. (1987). The effect of frontal eye field and superior colliculus lesions on saccadic latencies in the rhesus monkey. Journal of neurophysiology, 57(4), 1033-1049 [본문으로]
- Guitton, D., Buchtel, H. A., & Douglas, R. M. (1985). Frontal lobe lesions in man cause difficulties in suppressing reflexive glances and in generating goal-directed saccades. Experimental brain research, 58(3), 455-472 [본문으로]
- Shipp, S. (2003). The functional logic of cortico–pulvinar connections. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 358(1438), 1605-1624 [본문으로]
- Wurtz, R. H., & Goldberg, M. E. (1972). Activity of superior colliculus in behaving monkey. 3. Cells discharging before eye movements. Journal of Neurophysiology, 35(4), 575-58 [본문으로]
- Kelley, T. A., Serences, J. T., Giesbrecht, B., & Yantis, S. (2008). Cortical mechanisms for shifting and holding visuospatial attention. Cerebral cortex, 18(1), 114-125 [본문으로]
- Shomstein, S., & Yantis, S. (2006). Parietal cortex mediates voluntary control of spatial and nonspatial auditory attention. Journal of Neuroscience, 26(2), 435-439;Serences, J. T., Schwarzbach, J., Courtney, S. M., Golay, X., & Yantis, S. (2004). Control of object-based attention in human cortex. Cerebral cortex, 14(12), 1346-1357 [본문으로]
- Kelley, T. A., Serences, J. T., Giesbrecht, B., & Yantis, S. (2008). Cortical mechanisms for shifting and holding visuospatial attention. Cerebral cortex, 18(1), 114-125 [본문으로]
- Strayer, D. L., Drews, F. A., & Johnston, W. A. (2003). Cell phone-induced failures of visual attention during simulated driving. Journal of experimental psychology: Applied, 9(1), 23. [본문으로]
- Horrey, W. J., & Wickens, C. D. (2006). Examining the impact of cell phone conversations on driving using meta-analytic techniques. Human factors, 48(1), 196-205;Harbluk, J. L., Noy, Y. I., Trbovich, P. L., & Eizenman, M. (2007). An on-road assessment of cognitive distraction: Impacts on drivers’ visual behavior and braking performance. Accident Analysis & Prevention, 39(2), 372-379. [본문으로]
- Shomstein, S., & Yantis, S. (2004). Control of attention shifts between vision and audition in human cortex. Journal of neuroscience, 24(47), 10702-10706. [본문으로]
- Neisser, U., & Becklen, R. (1975). Selective looking: Attending to visually specified events. Cognitive psychology, 7(4), 480-494. [본문으로]
- Ophir, E., Nass, C., & Wagner, A. D. (2009). Cognitive control in media multitaskers. Proceedings of the National Academy of Sciences, 106(37), 15583-15587. [본문으로]
- Schneider, W., & Shiffrin, R. M. (1977). Controlled and automatic human information processing: I. Detection, search, and attention. Psychological review, 84(1), 1. [본문으로]
- Banich, M. T., & Compton, R. J. (2018). Cognitive neuroscience. Cambridge University Press [본문으로]
- Yantis, S., & Serences, J. T. (2003). Cortical mechanisms of space-based and object-based attentional control. Current opinion in neurobiology, 13(2), 187-193 [본문으로]
- O'Craven, K. M., Downing, P. E., & Kanwisher, N. (1999). fMRI evidence for objects as the units of attentional selection. Nature, 401(6753), 584-587 [본문으로]
- Mangun, G. R., Buonocore, M. H., Girelli, M., & Jha, A. P. (1998). ERP and fMRI measures of visual spatial selective attention. Human brain mapping, 6(5‐6), 383-389;Heinze, H. J., Mangun, G. R., Burchert, W., Hinrichs, H., Scholz, M., Münte, T. F., ... & Hillyard, S. A. (1994). Combined spatial and temporal imaging of brain activity during visual selective attention in humans. Nature, 372(6506), 543-546;Moran, J., & Desimone, R. (1985). Selective attention gates visual processing in the extrastriate cortex. Science, 229(4715), 782-784 [본문으로]
- Heinze, H. J., Luck, S. J., Mangun, G. R., & Hillyard, S. A. (1990). Visual event-related potentials index focused attention within bilateral stimulus arrays. I. Evidence for early selection. Electroencephalography and clinical neurophysiology, 75(6), 511-527 [본문으로]
- Corbetta, M., Miezin, F. M., Dobmeyer, S., Shulman, G. L., & Petersen, S. E. (1991). Selective and divided attention during visual discriminations of shape, color, and speed: functional anatomy by positron emission tomography. Journal of neuroscience, 11(8), 2383-2402 [본문으로]
- Anllo‐Vento, L., Luck, S. J., & Hillyard, S. A. (1998). Spatio‐temporal dynamics of attention to color: Evidence from human electrophysiology. Human brain mapping, 6(4), 216-238 [본문으로]
- Zhang, W., & Luck, S. J. (2009). Feature-based attention modulates feedforward visual processing. Nature neuroscience, 12(1), 24-25 [본문으로]
- Wojciulik, E., Kanwisher, N., & Driver, J. (1998). Covert visual attention modulates face-specific activity in the human fusiform gyrus: fMRI study. Journal of neurophysiology, 79(3), 1574-1578 [본문으로]
- Banich, M. T., & Compton, R. J. (2018). Cognitive neuroscience. Cambridge University Press [본문으로]
- Saalmann, Y. B., & Kastner, S. (2014). Neural mechanisms of spatial attention in the visual thalamus. In The Oxford Handbook of Attention (p. 399). OUP Oxford [본문으로]
- O'Connor, D. H., Fukui, M. M., Pinsk, M. A., & Kastner, S. (2002). Attention modulates responses in the human lateral geniculate nucleus. Nature neuroscience, 5(11), 1203-1209 [본문으로]
- Saalmann, Y. B., Pinsk, M. A., Wang, L., Li, X., & Kastner, S. (2012). The pulvinar regulates information transmission between cortical areas based on attention demands. science, 337(6095), 753-756 [본문으로]
- Strumpf, H., Mangun, G. R., Boehler, C. N., Stoppel, C., Schoenfeld, M. A., Heinze, H. J., & Hopf, J. M. (2013). The role of the pulvinar in distractor processing and visual search. Human brain mapping, 34(5), 1115-1132 [본문으로]
- Rafal, R. D., & Posner, M. I. (1987). Deficits in human visual spatial attention following thalamic lesions. Proceedings of the National Academy of Sciences, 84(20), 7349-7353;Snow, J. C., Allen, H. A., Rafal, R. D., & Humphreys, G. W. (2009). Impaired attentional selection following lesions to human pulvinar: evidence for homology between human and monkey. Proceedings of the National Academy of Sciences, 106(10), 4054-4059;Desimone, R., Wessinger, M., Thomas, L., & Schneider, W. (1990, January). Attentional control of visual perception: cortical and subcortical mechanisms. In Cold Spring Harbor symposia on quantitative biology (Vol. 55, pp. 963-971). Cold Spring Harbor Laboratory Press [본문으로]
- Antonio, R. D., Hanna, D., & Helena, C. C. (1980). Neglect following damage to frontal lobe or basal ganglia. Neuropsychologia, 18(2), 123-132 [본문으로]
- Bisiach, E., Geminiani, G., Berti, A., & Rusconi, M. L. (1990). Perceptual and premotor factors of unilateral neglect. Neurology, 40(8), 1278-1278 [본문으로]
- Isomura, Y., Ito, Y., Akazawa, T., Nambu, A., & Takada, M. (2003). Neural coding of “attention for action” and “response selection” in primate anterior cingulate cortex. Journal of Neuroscience, 23(22), 8002-8012 [본문으로]
- Cohen, M. X., Ridderinkhof, K. R., Haupt, S., Elger, C. E., & Fell, J. (2008). Medial frontal cortex and response conflict: evidence from human intracranial EEG and medial frontal cortex lesion. Brain research, 1238, 127-142 [본문으로]
- Wardak, C., Ibos, G., Duhamel, J. R., & Olivier, E. (2006). Contribution of the monkey frontal eye field to covert visual attention. Journal of Neuroscience, 26(16), 4228-4235 [본문으로]
- Kastner, S., & Pinsk, M. A. (2004). Visual attention as a multilevel selection process. Cognitive, Affective, & Behavioral Neuroscience, 4(4), 483-500 [본문으로]
- Gregoriou, G. G., Rossi, A. F., Ungerleider, L. G., & Desimone, R. (2014). Lesions of prefrontal cortex reduce attentional modulation of neuronal responses and synchrony in V4. Nature neuroscience, 17(7), 1003-1011 [본문으로]
- Baldauf, D., & Desimone, R. (2014). Neural mechanisms of object-based attention. Science, 344(6182), 424-427 [본문으로]
- Paus, T., Kalina, M., Patočková, L., Angerova, Y., Černý, R., Mečiř, P., ... & Krabec, P. (1991). Medial vs lateral frontal lobe lesions and differential impairment of central-gaze fixation maintenance in man. Brain, 114(5), 2051-2067 [본문으로]
- Coull, J. T., & Nobre, A. C. (1998). Where and when to pay attention: the neural systems for directing attention to spatial locations and to time intervals as revealed by both PET and fMRI. Journal of Neuroscience, 18(18), 7426-7435;Fink, G. R., Dolan, R. J., Halligan, P. W., Marshall, J. C., & Frith, C. D. (1997). Space-based and object-based visual attention: shared and specific neural domains. Brain: a journal of neurology, 120(11), 2013-2028 [본문으로]
- Knight, R. T., Scabini, D., Woods, D. L., & Clayworth, C. C. (1989). Contributions of temporal-parietal junction to the human auditory P3. Brain research, 502(1), 109-116 [본문으로]
- Kelley, T. A., Serences, J. T., Giesbrecht, B., & Yantis, S. (2008). Cortical mechanisms for shifting and holding visuospatial attention. Cerebral cortex, 18(1), 114-125;Nobre, A. C., Sebestyen, G. N., Gitelman, D. R., Mesulam, M. M., Frackowiak, R. S., & Frith, C. D. (1997). Functional localization of the system for visuospatial attention using positron emission tomography. Brain: a journal of neurology, 120(3), 515-53;Yantis, S., Schwarzbach, J., Serences, J. T., Carlson, R. L., Steinmetz, M. A., Pekar, J. J., & Courtney, S. M. (2002). Transient neural activity in human parietal cortex during spatial attention shifts. Nature neuroscience, 5(10), 995-1002 [본문으로]
- Corbetta, M., Patel, G., & Shulman, G. L. (2008). The reorienting system of the human brain: from environment to theory of mind. Neuron, 58(3), 306-324 [본문으로]
- Webb, T. W., Igelström, K. M., Schurger, A., & Graziano, M. S. (2016). Cortical networks involved in visual awareness independent of visual attention. Proceedings of the National Academy of Sciences, 113(48), 13923-13928 [본문으로]
- Corbetta, M., Patel, G., & Shulman, G. L. (2008). The reorienting system of the human brain: from environment to theory of mind. Neuron, 58(3), 306-324 [본문으로]
- Vallar, G., & Perani, D. (1986). The anatomy of unilateral neglect after right-hemisphere stroke lesions. A clinical/CT-scan correlation study in man. Neuropsychologia, 24(5), 609-622 [본문으로]
- Posner, M. I., Walker, J. A., Friedrich, F. J., & Rafal, R. D. (1984). Effects of parietal injury on covert orienting of attention. Journal of neuroscience, 4(7), 1863-1874 [본문으로]
- Ptak, R. (2012). The frontoparietal attention network of the human brain: action, saliency, and a priority map of the environment. The Neuroscientist, 18(5), 502-515 [본문으로]
- Gottlieb, J., Balan, P., Oristaglio, J., & Suzuki, M. (2009). Parietal control of attentional guidance: the significance of sensory, motivational and motor factors. Neurobiology of learning and memory, 91(2), 121-128 [본문으로]
- Vandenberghe, R., & Gillebert, C. R. (2009). Parcellation of parietal cortex: convergence between lesion-symptom mapping and mapping of the intact functioning brain. Behavioural brain research, 199(2), 171-182;Wojciulik, E., & Kanwisher, N. (1999). The generality of parietal involvement in visual attention. Neuron, 23(4), 747-764 [본문으로]
- Rensink, R. A. (2002). Change detection. Annual review of psychology, 53(1), 245-277;Simons, D. J., & Rensink, R. A. (2005). Change blindness: Past, present, and future. Trends in cognitive sciences, 9(1), 16-20. [본문으로]
- Simons, D. J., & Chabris, C. F. (1999). Gorillas in our midst: Sustained inattentional blindness for dynamic events. perception, 28(9), 1059-1074 [본문으로]
- Simons, D. J., & Levin, D. T. (1998). Failure to detect changes to people during a real-world interaction. Psychonomic Bulletin & Review, 5(4), 644-649. [본문으로]
- Furley, P., Memmert, D., & Heller, C. (2010). The dark side of visual awareness in sport: Inattentional blindness in a real-world basketball task. Attention, Perception, & Psychophysics, 72, 1327-1337. [본문으로]
- Rensink, R. A. (2002). Change detection. Annual review of psychology, 53(1), 245-277;Simons, D. J., & Rensink, R. A. (2005). Change blindness: Past, present, and future. Trends in cognitive sciences, 9(1), 16-20. [본문으로]
- Rensink, R. A., O'Regan, J. K., & Clark, J. J. (1997). To see or not to see: The need for attention to perceive changes in scenes. Psychological science, 8(5), 368-373. [본문으로]
- Hyman Jr, I. E., Boss, S. M., Wise, B. M., McKenzie, K. E., & Caggiano, J. M. (2010). Did you see the unicycling clown? Inattentional blindness while walking and talking on a cell phone. Applied Cognitive Psychology, 24(5), 597-607. [본문으로]
- Treisman, A. (1998). Feature binding, attention and object perception. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 353(1373), 1295-1306;Treisman, A. (2006). How the deployment of attention determines what we see. Visual cognition, 14(4-8), 411-443. [본문으로]
- Treisman, A., & Schmidt, H. (1982). Illusory conjunctions in the perception of objects. 1982, 14, 107-141. [본문으로]
- Treisman, A. M., & Gelade, G. (1980). A feature-integration theory of attention. Cognitive psychology, 12(1), 97-136;Treisman, A. (1998). Feature binding, attention and object perception. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 353(1373), 1295-1306;Treisman, A. (2006). How the deployment of attention determines what we see. Visual cognition, 14(4-8), 411-443;Treisman, A., & Schmidt, H. (1982). Illusory conjunctions in the perception of objects. 1982, 14, 107-141. [본문으로]
- Treisman, A., & Schmidt, H. (1982). Illusory conjunctions in the perception of objects. 1982, 14, 107-141;Treisman, A. (1998). Feature binding, attention and object perception. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 353(1373), 1295-1306 [본문으로]
- Robertson, L. C. (1999). What can spatial deficits teach us about feature binding and spatial maps?. Visual Cognition, 6(3-4), 409-430. [본문으로]
- Seymour, K., Clifford, C. W., Logothetis, N. K., & Bartels, A. (2010). Coding and binding of color and form in visual cortex. Cerebral cortex, 20(8), 1946-1954. [본문으로]
- Robertson, L. C., & Treisman, A. (1995). Parietal contributions to visual feature binding: evidence from a patient with bilateral lesions. Science, 269(5225), 853-855;Robertson, L. C. (2003). Binding, spatial attention and perceptual awareness. Nature Reviews Neuroscience, 4(2), 93-102. [본문으로]
- Braet, W., & Humphreys, G. W. (2009). The role of reentrant processes in feature binding: Evidence from neuropsychology and TMS on late onset illusory conjunctions. Visual Cognition, 17(1-2), 25-47. [본문으로]
- Friedman-Hill, S. R., Robertson, L. C., & Treisman, A. (1995). Parietal contributions to visual feature binding: evidence from a patient with bilateral lesions. Science, 269(5225), 853-855;Ashbridge, E., Walsh, V., & Cowey, A. (1997). Temporal aspects of visual search studied by transcranial magnetic stimulation. Neuropsychologia, 35(8), 1121-1131 [본문으로]
- Wardak, C., Olivier, E., & Duhamel, J. R. (2004). A deficit in covert attention after parietal cortex inactivation in the monkey. Neuron, 42(3), 501-508 [본문으로]
- Banich, M. T., & Compton, R. J. (2018). Cognitive neuroscience. Cambridge University Press [본문으로]
- Saper, C. B., Chou, T. C., & Scammell, T. E. (2001). The sleep switch: hypothalamic control of sleep and wakefulness. Trends in neurosciences, 24(12), 726-731;Edlow, B. L., Takahashi, E., Wu, O., Benner, T., Dai, G., Bu, L., ... & Folkerth, R. D. (2012). Neuroanatomic connectivity of the human ascending arousal system critical to consciousness and its disorders. Journal of Neuropathology & Experimental Neurology, 71(6), 531-546. [본문으로]
- Berridge, C. W., Schmeichel, B. E., & España, R. A. (2012). Noradrenergic modulation of wakefulness/arousal. Sleep medicine reviews, 16(2), 187-197;Sara, S. J. (2009). The locus coeruleus and noradrenergic modulation of cognition. Nature reviews neuroscience, 10(3), 211-223. [본문으로]
- Baker, J. L., Ryou, J. W., Wei, X. F., Butson, C. R., Schiff, N. D., & Purpura, K. P. (2016). Robust modulation of arousal regulation, performance, and frontostriatal activity through central thalamic deep brain stimulation in healthy nonhuman primates. Journal of neurophysiology. [본문으로]
- Sarter, M., Givens, B., & Bruno, J. P. (2001). The cognitive neuroscience of sustained attention: where top-down meets bottom-up. Brain research reviews, 35(2), 146-160;McGaughy, J., Kaiser, T., & Sarter, M. (1996). Behavioral vigilance following infusions of 192 IgG-saporin into the basal forebrain: selectivity of the behavioral impairment and relation to cortical AChE-positive fiber density. Behavioral neuroscience, 110(2), 247. [본문으로]
- Coull, J. T., Nobre, A. C., & Frith, C. D. (2001). The noradrenergic α2 agonist clonidine modulates behavioural and neuroanatomical correlates of human attentional orienting and alerting. Cerebral cortex, 11(1), 73-84. [본문으로]
- Paus, T., Zatorre, R. J., Hofle, N., Caramanos, Z., Gotman, J., Petrides, M., & Evans, A. C. (1997). Time-related changes in neural systems underlying attention and arousal during the performance of an auditory vigilance task. Journal of cognitive neuroscience, 9(3), 392-408. [본문으로]
- Portas, C. M., Rees, G., Howseman, A. M., Josephs, O., Turner, R., & Frith, C. D. (1998). A specific role for the thalamus in mediating the interaction of attention and arousal in humans. Journal of Neuroscience, 18(21), 8979-8989. [본문으로]
- HOWES, D., & BOLLER, F. (1975). Simple reaction time: evidence for focal impairment from lesions of the right hemisphere. Brain, 98(2), 317-332;Coslett, H. B., Bowers, D., & Heilman, K. M. (1987). Reduction in cerebral activation after right hemisphere stroke. Neurology, 37(6), 957-957. [본문으로]
- Yokoyama, K., Jennings, R., Ackles, P., Hood, P., & Boller, F. (1987). Lack of heart rate changes during an attention‐demanding task after right hemisphere lesions. Neurology, 37(4), 624-624. [본문으로]
- Sturm, W., Longoni, F., Fimm, B., Dietrich, T., Weis, S., Kemna, S., ... & Willmes, K. (2004). Network for auditory intrinsic alertness: a PET study. Neuropsychologia, 42(5), 563-568;Sturm, W., De Simone, A., Krause, B. J., Specht, K., Hesselmann, V., Radermacher, I., ... & Willmes, K. (1999). Functional anatomy of intrinsic alertness: evidencefor a fronto-parietal-thalamic-brainstem network in theright hemisphere. Neuropsychologia, 37(7), 797-805. [본문으로]
- Arruda, J. E., Walker, K. A., Weiler, M. D., & Valentino, D. A. (1999). Validation of a right hemisphere vigilance system as measured by principal component and factor analyzed quantitative electroencephalogram. International Journal of Psychophysiology, 32(2), 119-128. [본문으로]


