
| 일 | 월 | 화 | 수 | 목 | 금 | 토 |
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | ||
| 6 | 7 | 8 | 9 | 10 | 11 | 12 |
| 13 | 14 | 15 | 16 | 17 | 18 | 19 |
| 20 | 21 | 22 | 23 | 24 | 25 | 26 |
| 27 | 28 | 29 | 30 |
- 외측 슬상핵
- 테스토스테론렉스
- behavior modification
- 도덕발달단계론
- ctm
- 따뜻함주의
- #정신분석
- celtic knot
- collectivism
- 강화
- Psychology
- 행동주의
- 행동주의 치료
- 빅토리아 시대
- #산업및조직심리학
- 집단주의
- persuation
- 켈트 매듭
- 시각처리
- 개인주의
- criminal psychology
- 심리학
- sexdifference
- #정신역동
- memory
- individualism
- 연결주의
- 일주율
- #크립키
- heartism
- Today
- Total
지식저장고
눈에서 뇌까지: 시각의 흐름 본문
광할한 우주의 무를 누비며 돌아다니는 많은 빛들 중 일부는 우여곡절 끝에 우리의 눈으로 들어온다. 그러면 우리는 그 빛을 통해 빛이 거쳐온 광대한 세상을 비로소 볼 수 있게 된다. 이러한 두 우주적 존재의 만남은, 그러나 눈만 있어서는 안된다. 눈뿐만 아니라 눈으로 들어온 정보를 해석할 컴퓨터, 빛을 잘 여과해줄 렌즈, 빛에서 보이는 물체가 무엇인지 알게 해줄 장치, 시시각각 변하는 빛의 움직임을 포착할 동작감지기 등 무수한 장치가 있어야 제대로 빛을 볼 수 있다. 우주를 여행한 끝에 드디어 우리의 눈에 도착한 빛은 눈에서 시작하여 최종적으로 전두엽에 도달할 때까지 가공되고, 정련되면서 뇌 속을 흘러간다. 이러한 과정을 이해하면 시각장애인들을 돕고 뇌 지도를 파악하는데도 도움이 되지만, 한쪽 우주와 다른쪽 우주의 만남이 어떻게 진행되는지도 자세히 알수 있다.
빛에서 눈까지1

우리 눈으로 들어온 빛은 망막(retina)을 거쳐 색소층(pigmented epithelium)을 지나 광수용기(photoreceptor)에 도달한다. 광수용기는 간상체(rods)와 추상체(cones)로 나뉘는데 간상체는 명암을, 추상체는 색을 감지한다. 간상체는 12억개, 추상체는 500만개로 간상체가 더 많고, 눈의 초점이 위치한 중심와(fovea)에 추상체가 많이 분포하는 반면 간상체는 중심와 주변에 분포한다. 망막 내에 추상체가 많이 분포한 부분을 중심와라 하는데, 우리가 초점을 맞춘다는 말은 중심와에 상을 맺히게 한다는 말이다. 따라서 색은 초점을 맞출때 잘 보이는 반면, 밝기는 오히려 초점을 약간 비껴갈때 잘 보이는데, 이처럼 밝기는 잘 보이는 반면 색은 흐릿한 시야를 주변시(peripheral vision)이라 하며 한국군 군사교범에서도 어두울때는 목표물 옆에 초점을 맞추라고 교육하고 있다. 광수용기는 흔히 로돕신(rhodopsin, visual pigment molecules)이라고 부르는데, 간상체를 예를 들면 로돕신은 아래와 같이 생겼다.
로돕신은 오른쪽 가지에 붙은 retinal 부분과 몸통인 opsin으로 나눌수 있다. 로돕신은 이성질체로 cis형과 trans형을 가진다. 평소의 로돕신은 cis형인데, retinal이 광자에 반응하면 로돕신이 trans형으로 이성질화(isomerization)한다. 이성질화한 로돕신은 특정 효소를 활성화시키고 이 효소가 다른 효소와 반응해 연쇄반응(cascade)을 일으킨다. 눈에 들어온 빛의 단지 7%만이 로돕신과 반응하지만, 빛과 반응한 로돕신은 연쇄반응을 유발하면서 이를 증폭시킨다.
간상체와 추상체의 역치는 서로 다르다. 보통 추상체가 더 낮지만 암순응 상태가 되면 간상체의 역치가 더 낮다. 이들의 역치를 조절하는건 광수용기의 종류 뿐만이 아니다. 어느 광수용기든 가시광선 스펙트럼에서 중간에 가까울수록 역치가 낮다. 간상체는 500nm의 파장(초록-파랑의 경계)에서, 추상체는 560nm의 파장(연둣빛)에서 제일 역치가 낮다. 어두운 곳에서 암순응되면 푸른 빛이 더 밝게 보이는데(이를 퍼킨제 전이, purkinje shift라 한다) 이는 암순응 과정에서 일어난 rod-cone break로 인해 간상체가 우세해지기 때문에 일어난다. 여기서 고려해야 할 점은 추상체가 3가지 파장에 대응하는 광수용기를 가진다는 점이다. 빛의 3원색은 추상체에서 유래한다. 추상체는 각각 419nm, 532nm, 558nm에서 가장 민감한 3개의 광수용기를 가지는데 3원색이 이 파장대와 가깝다. 추상체는 이 3가지 수용기의 반응을 종합하여 받아들인 빛 자극이 어느 색인지 합성해낸다.
망막의 구조적 변화는 시각장애를 초래할 수 있다. 황반 변성(Macular degeneration)은 흔한 노인성 질환으로, 당뇨와 병행되는 이 병은 중심와와 주변이 파괴되면서 발생한다. 이 병에 걸리면 눈 중앙에 맹점이 생긴 것처럼 보이는데(이를 central vision이라 한다) 사실 그 자리는 중심와가 위치했던 자리다. 이와 반대의 경우는 retinitis pigmentosa라는 희귀성 유전질환으로, 이 병은 주로 간상체가 파괴된 뒤 중심와도 일부 공격받는다. 이 병을 앓는 환자들은 흐릿한 초점만 보이고,(이를 tunnel vision이라 한다) 안좋은 경우 중심와마저 파괴되어 시력을 잃는다.
연쇄반응을 일으킨 광수용기는 양극세포(bipolar cell)의 중계를 받아 뉴런인 신경절세포(ganglion cell, retinal ganglion cell, RGC)에 정보를 보내고 신경절세포에서 이를 전기신호로 변환한다. 보통 광수용기 9개당 양극세포가 6개, 신경절세포가 4개씩 붙기 때문에2 광수용기에서 내려오는 정보가 소수의 신경절세포로 수렴된다. 간상체 120개에서 발생한 정보가 하나의 뉴런에 수렴하는데 비해 추상체는 6개당 하나의 뉴런에 수렴하며 초점에서는 일대일로 대응한다. 그렇기 때문에 보통 분해능은 추상체가 더 좋고, 대신 간상체는 약한 자극을 지각하기 더 유리하다. 똑같은 빛 자극이 광수용기에 가해져도 추상체에서 수렴된 정보는 자극을 6배만 증폭시키지만 간상체는 120배로 증폭시켜서 약한 자극을 더 감지하기 쉽기 때문이다.
한편 아마크린 세포(amacrine cell)와 수평세포(horizontal cell)는 다른 세포와 달리 수평 방향으로 뻗어있는데 수평세포는 광수용기 사이를, 아마크린 세포는 신경절세포 사이를 중계하여 신경절세포의 수용장을 조절한다. 수용장(receptive field)은 한 세포가 정보를 받아들이는 시야의 부분으로, 대개 동물의 뉴런(신경절세포면 시신경)에 전극을 달고 망막 내 특정 위치에 빛을 비출때 전극이 발화하는지 여부로 수용장을 측정한다. 신경절세포의 수용장은 작으며(대개 중심와와 가까울수록 더 작다), m유형 뉴런은 시야 안의 운동에, p유형 뉴런은 모양에 관한 정보를 받아들인다. 수용장의 어디에 빛이 떨어지는지는 뉴런의 발화 여부와 관련되어 있는데, 보통 중심부는 흥분, 주변부는 억제를 일으킨다.(center-surround antagonism) 이런 구조는 각 물체의 가장자리를 인식하는데 유용하다. 이렇게 신경절세포의 수용장에 포착된 정보는 시신경(optic nerve)을 통해 외측 슬상핵(Lateral Geniculate Nucleus, LGN)으로 향한다.
암순응(dark adaption)
필자는 어릴때 어둠을 무서워했다. 아무것도 보이지 않아 어디서 귀신이 나올지 두려웠다. 그러다가 조금 시간이 지나면 모든 것들이, 비록 명확하진 않아도, 제대로 보였다. 그 때 보았던 은빛으로 물든 내 방, 내 집이 너무도 신기했다. 이런 경험은 누구나 한다. 이처럼 어둠 속에 있을때 시간이 지나면 어둠 속에서도 물체를 식별할 수 있는 현상을 암순응이라 한다. 암순응은 아주 자연스러운 반응으로, 처음 어둠에 직면하면 누구나 당황하지만, 주변자극이 사라졌기 때문에 오히려 광수용기의 민감도가 더 좋아진다. 지각심리학자들은 암순응을 연구하기 위해 피험자들을 어두운 방에 두고 시간이 지남에 따라 밝기의 역치를 측정했다. 피험자는 어두운 방에서 이미 보라고 했던 초점을 계속 응시하다가 초점 주변에서 빛이 보이는지 응답한다. 이는 초점에는 간상체가 없기 때문에 초점에 빛을 제시하면 높은 확률로 빛을 지각하지 못하기 때문이다.
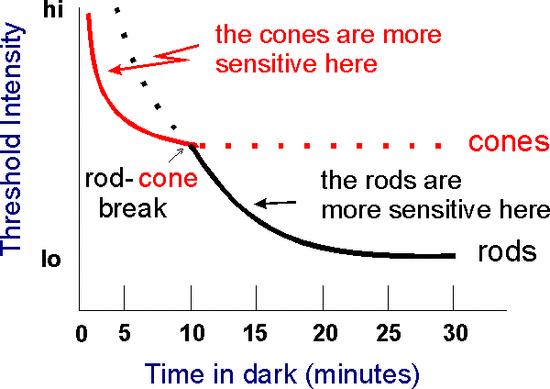
위 그래프는 암순응 실험에서 간상체와 추상체의 밝기 역치를 측정한 그래프이다. 위 그래프에서 보듯 암순응을 할때 광수용기의 민감도는 증가하는데(그래서 역치가 감소한다), 2개의 단계를 거쳐 증가한다. 첫번째 단계는 어두운 곳에 들어간 후 평균적으로 3-4분 동안으로, 이때 광수용기의 민감도는 계속 증가한다. 그러다가 두번째 단계가 7-10분까지 지속되는데, 이때는 민감도가 증가하는 폭이 작아진다. 보통 추상체가 간상체보다 빛에 민감하지만 이 단계에서 추상체의 민감도는 정체된다. 그래서 이 단계에서 간상체의 민감도가 추상체보다 높아지게 되는데 이를 rod-cone break라 한다. 이후 세번째 단계는 30분까지 지속되며 간상체의 민감도가 가파르게 증가하여 최고점에 달한다.
특이한 점은 간상체와 추상체가 암순응에 각각 다른 반응을 보인다. 이를 알기 위해 학자들은 간상체와 추상체의 역치를 따로 측정하는 방법을 고안해야 했다. 추상체의 경우는 쉽다. 초점에는 추상체만 있기 때문에 빛 자극을 제시하는 곳에 초점을 맞추게 하면 된다. 문제는 간상체다. 추상체가 없는 망막 표면은 없다. 이를 해결하기 위해 학자들은 다양한 방법을 사용한다. 피험자에게 선글라스를 씌워 추상체를 먹통으로 만들수도 있고, 아예 흑백만 구별가능한 단색 색맹(monochromat) 환자를 피험자로 사용하기도 한다. 이런 방법을 통해 위의 결과를 낼 수 있었다. 왜 암순응 현상이 발생할까? 아마 색소 형성시기와 관련있을지도 모른다. 우리가 시각을 유지하기 위해선 이성질화된 로돕신을 효소를 통해 다시 cis형으로 바꿔야 한다. 로돕신을 재형성하는데 추상체는 6분, 간상체는 30분이 걸린다. 각각 암순응에 필요한 시간과 비슷하다.
외측 억제(lateral inhibition)

위의 사진을 잘 보라. 아마 흰 선의 교차점 사이에 잠시동안 회색 점이 보일 것이다. 하지만 그 지점을 자세히 보면 회색 점은 사라진다. 이 착시를 hermann grid라 한다. 왜 이런 착시가 발생할까? 항상 그렇듯 착시는 우리의 심리를 더 잘 이해하게 해주는 지름길이다. 그리고 hermann grid는 그중에서도 망막의 수평선에 위치한 수평세포와 아마크린 세포의 중요성을 더 잘 보여준다.
위의 현상은 우리의 시각에서 수평선상의 세포들이 가지는 중요성을 보여준다. 이에 대한 지식은 투구게를 연구하면서 얻어졌다. 투구게는 원시적인 겹눈을 가진 살아있는 화석으로, 겹눈 구조는 광수용기가 크기 때문에 연구하기 쉽고 무척추동물이라 동물윤리도 크게 문제가 안된다. 그렇기 때문에 학자들은 시각을 연구하기 위해 투구게의 시각을 연구했는데 그러다가 특이한 현상을 발견했다. 투구게의 낱눈 하나에만 빛을 비추면 낱눈에 연결된 뉴런이 발화했다. 그런데 옆에 있는 낱눈도 같이 비추면 뉴런이 더 약하게 발화됬다. 옆의 낱눈에 더 강한 빛을 가하면 거의 평소 수준보다 더 약하게 발화했다. 이는 마치 낱눈이 서로를 억제하는 듯이 보인다.
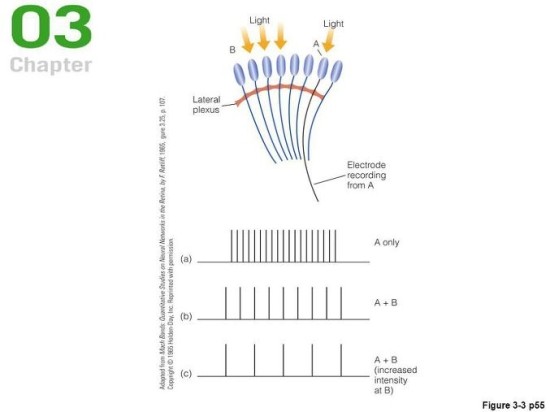
이런 현상은 수평선상의 세포들에 의해 일어난다. 수평으로 드러누워 광수용기나 신경절세포를 서로 연결하는 수평세포와 아마크린 세포는 광수용기의 정보가 신경절세포로 가는 사이에 간섭하여 서로 영향을 주게 한다. 이때 한 광수용기가 크게 발화하면 옆에 있는 다른 광수용기의 발화를 억제한다. 발화가 강할수록 억제도 강해진다. 이런 구조가 존재하는 이유는 이런 구조가 윤곽을 잘 구별하게 해주기 때문이다. Mach bands가 이를 잘 보여준다. mach bands는 연속적인 스펙트럼이나 색상을 보여줬는데도 사람들이 그 안에서 색을 구분시켜주는 윤곽이 있다고 믿는 착시이다. 무지개는 끊임이 없는 연속이지만 사람들은 5색이든 7색이든 무슨 색을 보고 경계선이 있다고 믿는다. 이 현상은 외측 억제 이론으로 매우 잘 설명된다. 수평차원의 세포가 인접한 광수용기들을 서로 억제하게 한다면 경계면 비스무리한 부분에서는 양 극단의 광수용기가 중간 지점의 광수용기에게 각각 다르게 작용한다. 이를 그림으로 보면 아래와 같다.
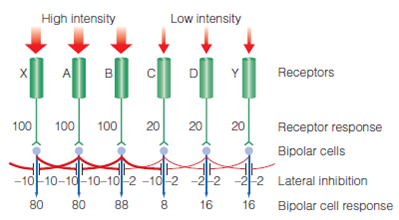
흑백 스펙트럼을 예로 들어보자. 아무리 스펙트럼이라도 흰색과 검은색의 구분이 모호한 지점은 존재한다. 그러한 지점에서 양 극단에 가까운 광수용기는 다른 광수용기와 차이가 없으나 중간에 위치한 광수용기는 다르다. 하얀 쪽의 광수용기는 하얀 쪽에서는 매우 강한 억제가 오지만(빛이 강해서) 검은쪽에서는 그렇지 않다. 그래서 극단 쪽의 광수용기와 달리 억제가 덜 되어 더 크게 발화한다. 반대로 검은 쪽의 광수용기는 검은 쪽에서는 약한 억제가 오지만 하얀 쪽에서는 매우 강한 억제가 온다. 그래서 극단 쪽과 달리 억제가 더 되어 더 작게 발화한다. 이렇게 되면 중간 지점의 광수용기는 다른 지점보다 더 밝거나 어두운, 더 유별난 발화를 보낸다. 이 둘의 차이는 둘이 차지하는 작은 지점이 유별나게 보이게 하고, 색대비로 인해 마치 흰색과 검은색 사이에 선이 있는 것처럼 보이게 한다. 이런 현상을 simultaneous contrast라 하는데, 이런 구조는 물체의 윤곽을 더 잘 구별하게 해주기 때문에 물체의 위치와 모양을 식별하는데 도움이 되어 동물의 생존을 돕는다.
hermann grid도 비슷하게 설명된다. hermann grid는 양극세포 수준에서 발생하는데 양극세포의 정보전달도 외측 억제의 영향을 받는다. hermann grid는 광수용기를 자극하는 흰색과 그렇지 않은 검정으로 구성되는데, 흰 선의 교차점을 중심으로 보면 주위 4방향의 양극세포가 모두 발화하게 된다. 발화한 4방향의 양극세포는 모두 인접 양극세포를 억제하고, 불행히도 사방이 둘러싸인 교차점 부분은 4개 세포의 억제를 모두 당하게 된다. 이로인해 교차점 부분은 포위 억제를 당해 회색으로 보이게 되고 우리가 초점을 교차점에 두어 시야를 더 조밀하게 만들면(투입되는 광수용기를 늘리면) 회색 점이 사라진다.
외측 억제는 not 회로와 단순 입력 회로를 모두 가진 뉴런의 특성을 통해 가능하다. 모든 뉴런은 하나의 논리 연산자로, 이는 망막도 다름이 없다. 보통 광수용기에서 신경절세포로 이어지는 흐름은 단순한 입력 회로다. 광수용기가 발화하면 신호가 그대로 수렴되어 신경절세포로 이어진다. 그러나 수평 차원에서는 약간 다른 일이 일어난다. 수평 차원의 세포를 통해 만들어진 광수용기간 회로는 흥분성이 아닌 억제성일 수도 있다. 이런 경우 한 광수용기의 발화는 신경 회로를 타고 흘러가 다른 광수용기를 억제한다. 물론 신경절세포는 자기가 담당하는 광수용기와 가장 강하게 연결되기 때문에 타 광수용기의 억제 신호보다는 자기 광수용기의 흥분 신호에 더 민감하다. 하지만 타 광수용기가 너무 강하게 발화하면 이는 무너진다. 결과적으로 다른 광수용기와 비교하여 한 광수용기가 중간 정도로 활성화될때 가장 큰 자극이 신경절세포로 보내진다.
외측 억제는 수많은 착시를 설명한다. 그러나 다는 아니다. 여기에는 예외가 있다. 대표적인 예는 benary cross와 white's illusion이 있다.


위 그림의 두개의 삼각형이 보이는가? 둘의 색깔이 같은가? 사실 둘의 색깔은 같지만 사람들은 왼쪽 삼각형이 더 밝은 듯한 착시를 느낀다. 외측 억제 이론을 따른다면, 둘 다 같은 정도로 광수용기가 억제되기 때문에 둘이 같은 색깔로 보여야 한다. 아래쪽은 어떤가? A에 어렴풋이 보이는 막대와 B에 어렴풋이 보이는 막대의 색이 같은가? 외측 억제 이론을 따르면 A가 더 밝아야 하지만 실제로는 반대다. 이 두가지 현상은 아직까지도 미스터리로 남아있다. 학자들이 세운 정설은 배경과 관련되어 있다. 가설에 따르면 benary cross는 왼쪽 삼각형은 하얀 배경이 검은 십자 위에 묻은 형태로, 오른쪽은 검은 십자가 하얀 배경 위에 묻은 형태로 인식되기 때문에 일어난다. white's illusion은 A가 검은 선 뒤에, B는 검은 선 앞에 있다고 인식되어 일어난다. 이 둘은 모두 가설이라는 점과 외측 억제 이상의 차원으로 착시를 설명한다는 공통점이 있다. 실제로 뮐러-라이어 착시같은 많은 착시는 대뇌 수준에서 일어난다. 당신이 착시에 관심이 많다면 당신의 눈뿐만 아니라 뒤에 있는 뇌까지 뒤져야 할 것이다.
1.2.눈에서 뇌까지3
눈에서 빠져나온 정보는 외측 슬상핵에 도달하기 전 시교차(chiasm, optic chiasma)에서 경로가 엇갈린다. 여기서 좌측 시야는 모두 좌측으로, 우측 시야는 모두 우측으로 향한다.(이 과정을 거치기 전 단계를 optic nerve, 후 단계를 optic track이라고 한다.) 이렇게 교차된 정보는 시각속(optic tract)을 통해 외측 슬상핵으로 향해서 간단한 가공을 거친다.
외측 슬상핵은 무릎처럼 생긴 기관으로 6개 세포층으로 구성되어 있다. 2,3,5층은 눈에서 나온 시신경과 바로 연결되고 1,4,6층은 반대쪽 시신경과 연결된다. 이 층들 중 1,2층을 구성하는 거대세포층(Magnocelluar)에서는 m유형 뉴런에서 온 운동 정보를, 3,4,5,6층을 구성하는 Parvocellular 층에서는 p유형에서 온 모양 정보를 처리하고, 과립성 세포층(koniocellular)에서는 m도 p도 아닌 신경절세포로부터 받은 색채 정보를 처리한다. 처리하는 정보유형에 따라 LGN의 층들은 magno 채널과 parvo 채널로 분류되는데, magno 채널은 운동 정보를 처리하고, parvo 채널은 과립성 세포층과 parvocellular 층을 포함하여 모양, 색체, 질감, 깊이 등을 처리한다.
이러한 분류는 대뇌 수준까지 이어진다. 아직 이 단계에서 수용장의 크기는 변함이 없으며, 보통 도넛 모양을 한 수용장에 점 자극만 가해져도 반응한다. 외측 슬상핵은 처리한 정보 일부를 편도체로 보내고, 모든 정보를 후두엽(occipital lobe)4의 1차 시각피질(primary visual cortex, vision cortex, V1)로 보낸다.
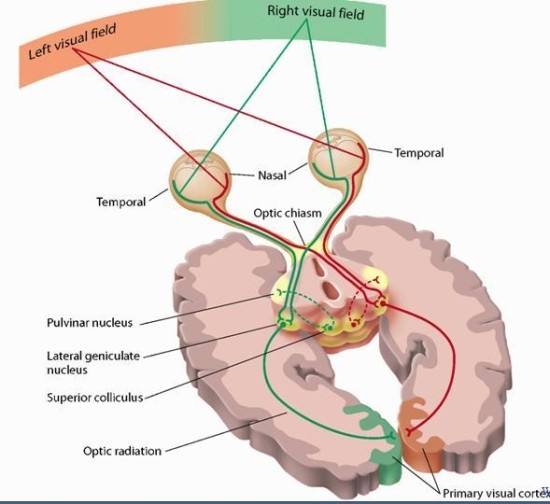
망막에서 1차 시각피질에 이를때까지 시각 정보는 큰 변화를 겪지 않는데 이를 연결대응(retinotopy)이라 부른다. 연결대응은 시야와 수용장을 위상적으로 보존한다. 즉 신경절세포의 수용장은 모양과 크기는 바뀔지언정 위상수학적 특성은 바뀌지 않는다. 그래서 V1의 수용장을 늘이고 조이고 줄이면 신경절세포의 수용장을 만들 수 있다. 시야에서 각 지점의 상대적 위치도 위상적으로 보존되어 V1까지 간다.(이를 retinotopic mapping이라 한다) 실제로 연구자들이 고양이의 V1에 전극을 수평으로 꽃고 앞에서 4가지 색의 사각형 배열을 움직이며 보여주자 고양이의 blob이 각기 다르게 반응했는데, 이 패턴이 망막상의 그것과 유사했다. 다만 V1에서의 수용장은 사영기하학적으로는 망막과 차이가 있다. 초점은 망막 수용장의 0.01%만 차지하지만 V1에서는 8-10%를 차지한다. 그래서 우리의 초점에 들어온 정보는 더 잘 지각되고 우리의 주의를 끈다. 다만 뇌의 상위 영역에서 이를 수정하기 때문에 우리는 실제 지각된 것보다 작은 초점 영역과 선명한 주변시를 의식적으로 느끼게 된다. 과학자들은 연결대응 현상을 알아보기 위해 다소 끔찍한 실험을 고안했다.
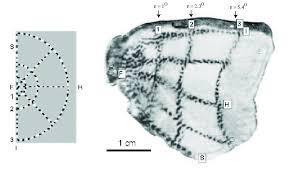
연구자들은 원숭이를 묶어놓고 방사성 동위원소를 투여한 링거를 투여하면서 왼쪽의 자극만 보여줬다. 원숭이는 죽을때까지 자유를 박탈당한채 형틀에 묶여 시각 자극만 계속 보아야 했다. 긴 시간이 지나면 자극의 선을 지각하는 피질 영역이 피로하게 되고 방사성 동위원소를 통해 검은색을 띤다. 때가 충분해지면 연구자들은 원숭이를 죽여 뇌를 적출한다. 그리고 뇌를 얇게 저며서 뉴런의 패턴을 분석한다. 실험결과 시각피질의 활동은 보는바와 같이 실제 자극과 유사했고 연결대응을 증명했다. 또한 실제 자극에 비해 초점이 위치하는 부위가 확장된 모습을 보여주어, 초점이 가지는 중요성을 증명하였다. 그리고 연구자들은 의도하지 않았지만, 인간이 자기들의 호기심을 위해 어떻게 한 생명체를 잔인하게 학대하고 죽일수 있는지도 증명한 듯 하다.
연결대응된 정보들은 V1에 이르러서 더 복잡하고 정교하게 처리된다. 후측 후복합(posterial occipital complex)을 구성하는 시각피질에서 시각정보를 가장 먼저 받는 선조피질(striate cortex)에는 다양한 수용장을 지닌 다양한 뉴런이 있다. 어떤 뉴런은 특정 방위에, 어떤 뉴런은 특정 운동에 반응한다.5 simple cortical 세포는 길이를 지각한다. 이 세포의 수용장은 LGN에서 올라온 세포들의 수용장 여러개를 한줄로 겹쳐놓은 모양인데, 이런 구조는 특정 길이나 방향을 잘 지각하게 해준다. 방향을 지각하는 일은 가장 중요한 일 중 하나로, 사람은 어느 물체가 수직방향인지 수평방향인지를 중요하게 여긴다. 실제 연구결과 인간은 수평이거나 수직인 방향을 다른 방향보다 더 잘 지각했으며(이를 경사 효과, oblique effect라 한다) 어떤 방향이 수직이나 수평에서 얼마나 먼지 꽤 정확하게 측정한다. 이는 인간의 본능으로 보인다.
complex cell(복합세포)은 simple cortical 세포와 비슷하지만, 특정 방향으로의 이동도 지각할 수 있다. end-stopped 세포는 hypercomplex cell(과복합세포)이라고도 불리는데, 이 세포는 움직이는 특정 길이와 각도를 가진 물체에만 반응한다. 시각피질은 이처럼 같가지 다양한 뉴런들이 폭이 넓은 선이나 시야 주변의 선 등 특정 자극에만 특화되어 반응한다.6 그리고 이런 뉴런들은 서로 모여 기둥(column)을 이루어 여러 정보를 처리한다. 뉴런들의 중심에 위치한 신경기둥 블럽(blob)은 과립성 세포층에서 올라온 색 정보를 처리하는 조직으로 영장류보다는 고양이가 더 발달해 있다. location column은 위치 정보를 처리하는데 연결대응이 여기서도 이어진다. 그리고 orientation column은 특정 방향에 반응하는 뉴런의 정보를 통합하는데 두께가 1mm도 되지 않는 이 기둥에서 방향에 대한 모든 정보가 처리된다. ocular dominance column은 특정 눈의 정보만 처리하는 0.25-0.5mm의 기둥으로 왼눈과 오른눈을 구별한다. 이외에도 많은 신경기둥들이 시각정보를 통합한다. 신경기둥은 복잡한 시각정보들을 좀더 간단하게 정리하지만, 신경기둥 자체는 꽤나 복잡한 구조를 가졌다.
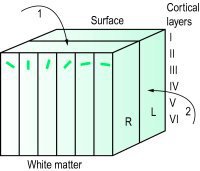
신경기둥은 대개 hypercolumn이라 불리는 더 큰 구조의 일부인데 이 구조는 3차원으로 이해해야 잘 이해된다. 맨 위 표면은 location cell이 분포하여 연결대응을 유지하며 LGN에서 올라온 시각정보를 그대로 투사한다. 불행한 원숭이에서 봤듯이 시야에 비친 모습 거의 그대로의 시각정보가 윗표면에 투사되면, 아래에 있는 신경기둥들이 정보를 처리한다. 여기서 주의할 점은 R과 L로, 각각 왼눈과 오른눈만 담당하는 ocular dominance column을 나타낸다. hypercolumn의 중심에는 블럽이 자리하여 색채 정보를 통합하는데 이 구조는 영장류에서는 잘 드러나지 않는다.
complex cell에 관해 떠오르는 의문은 complex cell이 운동을 지각할 수 있는지이다. 이 문제를 aperture problem이라 하는데, 이게 왜 문제가 되는지는 아래 사진의 위쪽 이미지를 보자.
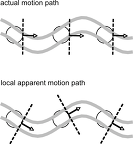
위의 동그라미는 뉴런의 수용장, 곡선은 선분의 움직임을 나타낸다. 저 그림에서 선분은 구불구불하게 움직인다. 하지만 선분이 직선으로 움직일 수도 있다. 그리고 저 작은 수용장 안에서만 보면 선이 구불구불하게 움직이는지 똑바로 움직이는지 알수 없다. 수용장 안에서는 선이 왼쪽에서 오른쪽으로 지나가는 것만 볼수 있기 때문이다. 학자들은 뇌의 상위 영역에서 각 complex cell이 보낸 정보를 모아 전체적으로 구성하여 움직임을 지각한다고 보며, 측두엽의 medial temporal cortex에서 이같은 일이 일어난다.
이처럼 후두엽으로 들어온 시각정보는 여러 뉴런과 신경기둥의 협업을 통해 하나의 통합된 상을 구현된다. 눈에 이상이 없음에도 시각에 장애를 가친 실인증(agnosia)7 환자들 중에는 대상의 방위나 촉감 등은 인지하지만 대상의 통합된 모습은 인지하지 못하는 환자가 있는데, 이를 통각 실인증(apperceptive agnosia)이라 부르며 시각피질이 망가진 결과로 보고 있다. V1 외에도 대상의 색, 질감, 길이를 처리하는 V2, 색을 깊이있게 처리하는 V4 등 30-50개에 달하는 다양한 피질이 후두엽을 이루고 있으며,8여기에 몇몇 측두엽 영역도 합세하여 시각정보를 처리한다. 이렇게 처리된 시각정보는 체계화되어 다른 뇌부위로 보내지는데, 보통 거대세포층에서 처리되는 운동, 위치 등의 정보는 측두엽의 medial temporal cortex를 거쳐 두정엽으로 보내지고, parvocelular 층에서 처리되는 모양, 개념 등은 측두엽으로 보내진다.9
각 정보를 처리하는 뉴런은 원래 그런 기능을 하도록 유전자 수준에서 설계되어 있지만 그렇다고 환경이 중요하지 않은건 아니다. 고양이를 대상으로 한 실험에서, 과학자들은 갓 태어난 새끼고양이에게 깔때기를 씌우고 가로줄무늬로 가득한 방에 살게 하였다. 그리고 고양이가 어른이 되자 시각피질에 전극을 꽃고 활성화 패턴을 관찰하였다. 관찰결과 다른 고양이에 비해 이 고양이는 세로방향에 반응하는 뉴런 활동이 보이지 않았다. 세로줄무늬 방에 살게 한 고양이는 반대로 가로방향에 반응하는 뉴런 활동이 보이지 않았다. 이는 고양이가 가로세로 모두 볼 수 있는 뉴런을 가지고 태어났음에도 불구하고, 환경 자극을 통해 피드백을 받지 못해 해당 뉴런이 소거되었기 때문이다. 실제로 동물실험 결과 특정 시기동안 시각을 사용하지 못하면 동물은 눈의 건강과 상관없이 시각을 영구히 잃어버린다. 유전적으로 프로그래밍된 뉴런도 환경이 같이 있어야 제대로 발달할 수 있다.
배측 경로(dorsal pathway)와 복측 경로(ventral pathway)를 통해 후두엽의 정보는 두정엽과 측두엽으로 보내진다.10 이러한 경로는 미슈킨(Mishkin)과 동료들의 실험에서 발견되었다. 미슈킨은 어떤 원숭이는 두정엽의 절반을, 어떤 원숭이는 측두엽의 절반을 잘라내고 두가지 과제를 시켰다. 하나는 object discrimination task인데, 이 과제에서 원숭이는 두 물체를 보고 학습한 물체를 집어야 한다. 이 과제는 물체를 정확하게 인지하고 골라내는 능력이 필요하다. 다른 하나는 Landmark discrimination task인데, 이 과제에서 원숭이는 실린더 옆에 놓인 물체를 집어야 한다. 이 과제는 물체의 위치를 정확하게 인지하고 정교하게 집어내는 능력이 필요하다. 실험결과 두정엽이 손상된 원숭이는 landmark discrimination task를 하지 못했고 측두엽이 손상된 원숭이는 object discrimination task를 하지 못했다. 이는 두정엽이 물체의 위치와 운동 정보를 처리하고 측두엽이 물체를 재인한다는 사실을 보여준다. 일산화탄소 노출로 측두엽이 손상된 인간 환자 DF11도 카드를 리더기 비스무리한 구멍에 꽃을 순 있었지만, 연구자가 방향을 맞춰서 꽃으라는 지시사항을 추가하자 방향을 몰라 안절부절했다.
후두엽에서 두정엽으로 향하는 경로는 위치에 따라서 배측 경로(dorsal pathway)라 부르거나, 어떻게 하는지 혹은 어디에 있는지에 관한 정보를 처리한다고 how pathway12 혹은 where pathway13라 부른다. 측두엽으로 향하는 경로도 비슷하게 복측 경로(ventral pathway)라 부르거나 물체가 무엇인지에 대한 정보를 처리한다고 what pathway라 부른다. 그러나 일부 운동과 관련된 정보는 측두엽으로 보내진다. 이 두 경로가 서로 상호작용하는지는 아직 논쟁중이다. milner와 goodale은 배측 경로가 운동과 연관된다고 action pathway로, 복측 경로는 물체 지각과 관련된다고 perception pathway로 명명하였다.
두정엽에서는 보통 무의식적으로 정보가 처리되고 실질적인 운동에 초점이 맞춰지는 반면 측두엽에서는 정보가 의식 수준에 가깝게 처리되고, 물체의 재인이 이루어진다.14 처음 도착한 정보는 먼저 측두엽 가장 뒤편의 외측 후복합(lateral occipital complex, LOC)에서 기본적으로 처리되고, 이어 IT 피질(Inferior temporal cortex)에로 이동한 뒤 앞면으로 순차적으로 진행하면서 더 깊이있게 처리된다. 여기서 시각정보는 기존의 기억과 연결되면서 비로소 뇌는 대상이 어떤 물체인지 파악한다.
ventral pathway나 dorsal pathway가 망가지는 방식에 따라 다양한 실인증이 나타날 수 있다. 연합 실인증(associative agnosia)을 앓는 환자들은 통합된 지각이 가능하여 본 물체를 그리는게 가능하면서도 정작 자신이 그린게 무엇인지, 자신이 보는게 무엇인지 알지 못하는데, 보통 외측 후복합이 손상되어 복측 경로가 파괴되면 이러한 일이 생겨난다. 손상 정도가 덜한 경우 큼지막한 재인은 가능한 경우도 있고(고양이는 몰라도 털 달린 동물인건 안다), 시각 경로만 망가졌기 때문에 촉각, 청각 등을 통한 지각은 통각 실인증과 마찬가지로 가능하다. 동시실인증(simultanagnosia)은 2가지 이상의 시각 속성을 통합하지 못하는 실인증인데, 동시실인증 환자는 물체의 색을 지각할 수는 있으나 그것이 해당 물체의 색이라고 지각할 수 없다. 동시실인증은 dorsal pathway의 손상에 의한 것으로 보인다.15
특이한 점은 측두엽이 물체를 재인하는 방식인데, 어떤 물체는 그 물체를 지각하는 부위가 따로 있다.16 이를 과학에선 모듈화라고 한다. 이런 신기한 사실은 원숭이를 연구하던 부주의한 연구자가 원숭이 앞에서 손을 흔들자 원숭이 뇌의 특정 부위가 강하게 반응한 사건에서 발견되었다. 일례로 원숭이의 IT 피질에서는 오직 막대사탕(lollipop) 모양만 지각하는 뉴런이 발견되었다. IT 피질은 이외에도 수많은 특정 자극들만 지각하는 모듈이 있고, IT 피질과 인접한 후두엽의 Extrastriate Body Area(EBA)에서는 전체 몸과 신체 일부만 지각한다.
비슷하게 우뇌의 방추상회(fushiform face area,FFA)는 얼굴 정보를 주로 처리하는데, 이들의 수용장은 거의 시야 전체를 커버할 수 있을 정도로 크고, 그 안에서 오로지 얼굴에만 반응한다. 다른 영역도 마찬가지인데 역시 우뇌의 PPA(parahippocampal place area)도 역시 큰 수용장을 가졌으며 장소 관련 정보에만 반응한다. 창살(grating) 형태의 자극을 selective adaptation한 연구에 따르면, 한 방향의 창틀만 계속 보여주면 뉴런이 해당 방향의 창틀 자극에는 습관화되는데 비해 다른 방향의 창틀에는 습관화되지 않았다. 이러한 습관화는 방향뿐만 아니라 크기에서도 일어난다. 과학자들은 특정 크기만 지각하는 뉴런을 확인하기 위해 아래와 같은 착시를 고안했다.

a에서 두 창살모양 사이에 있는 막대를 응시하라. 막대의 좌우 끝으로 눈동자를 마구 움직여라. 2분동안 그러고 있으면 높은 확률로 피곤해진다. 사실 이 피곤함이 제일 중요하다. 2분동안 시간을 허비했으면 이제 b의 막대를 보라. 아마 위의 창살은 더 굵게, 아래 창살은 더 얇게 보일 것이다. 이런 현상은 당신이 피곤하기 때문에 일어난다. 당신이 2분동안 a를 쳐다보는 동안 당신 뇌 속의 뉴런은 각기 다른 상황에 처한다. 시야 윗쪽에 수용장을 가진 뉴런 중 얇은 크기에 특화된 뉴런은 계속 얇은 선만 쳐다보느라 피곤해진다. 마찬가지로 아래쪽의 뉴런 중 굵은 크기에 특화된 뉴런도 피곤해진다. 이들은 2분동안 너무 힘들게 일했기 때문에 더 일할 여력이 없다. 그래서 당신이 b로 시야를 옮기든 말든 태업을 한다. 그렇게 되면 윗쪽에는 굵은 크기를 담당하는 뉴런이, 아래쪽은 얇은 크기를 담당하는 뉴런이 홀로 남아 일하게 되고, 결국 우리 눈에는 위는 굵게, 아래는 얇게 보인다. 이런 실험을 통해 학자들은 크기에 특화된 뉴런도 존재함을 밝혀냈다.
측두엽과 두정엽에서 처리된 시각정보는 마침내 뇌 곳곳의 필요한 곳에 전해진다. 그중 가장 중요한 곳은 시각정보를 해석하는 전두엽, 특히 PFC다. PFC는 두정엽과 측두엽이 처리한 시각정보를 바탕으로 어떤 현상을 해석하거나 추론하고 그에 따라 적절하게 행동한다. 이를 연구한 어떤 실험에서는 원숭이에게 개나 고양이 사진을 하나 보여주고 이후에 개와 고양이를 적절히 섞은 사진을 보여주었다. 원숭이는 사진이 개에 가까운지 고양이에 가까운지 잘 구분해야 먹이를 얻을 수 있었다. 원숭이의 뇌를 측정한 결과 구분하기 전 샘플 이미지를 볼때는 IT 피질이 활동했지만, 샘플 이미지를 본 후와 직접 사진을 개나 고양이로 구분할 시점에는 PFC가 활동했다. 이는 시각정보를 처리하는건 측두엽일지라도 이를 기초로 생각하고 판단하는건 전두엽임을 보여준다. 차를 운전하다 노란 신호등을 보았을때 불이 노란색임을 인지하는건 측두엽이지만, 노란색을 빨간불의 예비신호로 인식하고 차를 세울지 아니면 빨간불까지 얼마 남지 않은 시간으로 인식하고 엑셀을 밟을지는 PFC가 결정한다.
최근 연구자들은 위에서 말한 경로 외에도 또다른 경로가 있음을 발견했다. 새로이 발견된 사실에 따르면 LGN은 일부 정보를 시상에 위치한 superior colliculus(SC)로 보낸다. 그러면 superior colliculus는 다시 정보를 두정엽으로 보내는데 이 정보가 무엇인지는 아직 밝혀지지 않았다. 쥐의 경우 SC에서 방향 탐지에 사용되는 것과 유사한 세포구조가 발견되었다.17 인간도 그러한지는 미지수다.
이 분야의 주요 연구자로는 故데이비드 휴벨(David Hubel), 토르스텐 비셀(Wissel), 반 에센(Van essen), 그리고 마츠모토(Matsumoto)가 있다. 휴벨과 위젤은 방위나 모양에 반응하는 뉴런을 찾아낸 공로18로 노벨상을 수상했다. 마츠모토는 sparse coding과 population coding 사이의 논쟁에서 주요 역할을 맡고 있다.
- Gerrig,'심리학과 삶 20',이종한 외 5인 역,시그마프레스,2016,p77 [본문으로]
- 망막에는 1억 2600만개의 광수용기와 100만개의 신경절세포가 있다. [본문으로]
- Gerrig,'심리학과 삶 20',이종한 외 5인 역,시그마프레스,2016,p78;Schacter 외 2인,'심리학 입문(2)',민경환 외 8인 역,시그마프레스,pp112-113 [본문으로]
- Zeki, S. (2001). Localization and globalization in conscious vision. Annual review of neuroscience, 24(1), 57-86. [본문으로]
- Hubel, D. H., & Wiesel, T. N. (1962). Receptive fields, binocular interaction and functional architecture in the cat's visual cortex. The Journal of physiology, 160(1), 106;Hubel, D. H., & Wiesel, T. N. (1998). Early exploration of the visual cortex. Neuron, 20(3), 401-412;Hubel, D. H. (1995). Eye, brain, and vision. Scientific American Library/Scientific American Books [본문으로]
- Livingstone, M., & Hubel, D. (1988). Segregation of form, color, movement, and depth: anatomy, physiology, and perception. Science, 240(4853), 740-749 [본문으로]
- Goodale, M. A., & Milner, A. D. (1992). Separate visual pathways for perception and action;Goodale, M. A., & Milner, A. D. (2004). Plans for action. Behavioral and Brain Sciences, 27(1), 37. [본문으로]
- Orban, G. A., Van Essen, D., & Vanduffel, W. (2004). Comparative mapping of higher visual areas in monkeys and humans. Trends in cognitive sciences, 8(7), 315-324;Van Essen, D. C., Anderson, C. H., & Felleman, D. J. (1992). Information processing in the primate visual system: an integrated systems perspective. Science, 255(5043), 419-423. [본문으로]
- Tanaka, K. (1996). Inferotemporal cortex and object vision. Annual review of neuroscience, 19(1), 109-139. [본문으로]
- Konen, C. S., & Kastner, S. (2008). Two hierarchically organized neural systems for object information in human visual cortex. Nature neuroscience, 11(2), 224-231. [본문으로]
- Goodale, M. A., Milner, A. D., Jakobson, L. S., & Carey, D. P. (1991). A neurological dissociation between perceiving objects and grasping them. Nature, 349(6305), 154-156. [본문으로]
- Milner, A. D., & Goodale, M. A. (1995). Oxford psychology series, No. 27. The visual brain in action. [본문으로]
- Ungerleider, L. G., & Mishkin, M. (1982). Two cortical visual systems. Analysis of visual behavior. Ingle DJ, Goodale MA, Mansfield RJW. [본문으로]
- Martin, A. (2007). The representation of object concepts in the brain. Annu. Rev. Psychol., 58, 25-45 [본문으로]
- Coslett, H. B., & Lie, E. (2008). Simultanagnosia: Effects of semantic category and repetition blindness. Neuropsychologia, 46(7), 1853-1863;Coslett, H. B., & Lie, G. (2008). Simultanagnosia: when a rose is not red. Journal of Cognitive Neuroscience, 20(1), 36-48. [본문으로]
- Mahon, B. Z., & Caramazza, A. (2011). What drives the organization of object knowledge in the brain?. Trends in cognitive sciences, 15(3), 97-103. [본문으로]
- Feinberg, E. H., & Meister, M. (2015). Orientation columns in the mouse superior colliculus. Nature, 519(7542), 229 [본문으로]
- Hubel, D. H., & Wiesel, T. N. (1962). Receptive fields, binocular interaction and functional architecture in the cat's visual cortex. The Journal of physiology, 160(1), 106;Hubel, D. H. (1995). Eye, brain, and vision. Scientific American Library/Scientific American Books [본문으로]

